

 下载:
下载:


 下载:
下载:

-
槟榔为棕榈科植物槟榔的干燥成熟种子,居“四大南药”之首,入药最早记录在《李当之药录》中[1],具有行气利水,消积杀虫,截疟等功效,用于治疗寄生虫病、关节炎、青光眼、水肿、脚气、疟疾等疾病[2]。槟榔主要含生物碱、多酚、多糖、脂肪酸、鞣质、氨基酸、三萜类、矿物质、槟榔红色素、粗纤维、油脂、维生素等多种物质[3-4]。药理研究表明其具有抗氧化、抗抑郁、抗炎、抗寄生虫、抗老化、清除自由基、修护神经系统、抗疲劳、改善胃肠功能、治疗糖尿病、抑菌等作用[5],文献报道槟榔的抗氧化作用与其含有的多酚类物质有关[6-12]。
课题组前期的研究发现,槟榔醇提取物能显著增强H9c2细胞的耐缺氧能力[13],进行高原实地研究发现,槟榔多酚能够显著改善高原实地缺氧大鼠的血气指标,能提高其肝、肺、心组织中谷胱甘肽、超氧化物歧化酶活性,降低其肝、肺、心组织中丙二醛的含量,槟榔多酚提取物的抗缺氧作用与消除过量的氧自由基,减轻脂质过氧化和氧化应激损伤有关[14-15]。同时,课题组进一步研究还发现,槟榔多酚提取物能预防高原肺水肿的发生,该作用与其减轻肺水肿、维持肺泡毛细血管通透性、改善氧化应激损伤及病理损伤有关[16]。由此可见,将槟榔多酚提取物开发为新的抗高原缺氧药物具有很大的潜力。槟榔多酚成分复杂,文献报道槟榔多酚中含有儿茶素、表儿茶素、原儿茶酸、没食子酸等,另外药理研究表明,儿茶素、表儿茶素、原儿茶酸具有抗氧化、抗炎等作用[17-21],基于此,本研究采用紫外分光光度法对槟榔多酚提取物中总多酚的含量进行测定;并建立槟榔多酚提取物中儿茶素类成分的HPLC测定方法,以期为槟榔多酚提取物的质量控制和标准制定提供实验依据。
-
槟榔由联勤保障部队第九四〇医院药剂科中药房提供,没食子酸(上海源叶生物科技有限公司,批号Y19M8C36143,质量分数≥98%)、儿茶素(中国食品药品检定研究院,批号110877-201604,质量分数99.2%)、表儿茶素(上海阿拉丁生化科技股份有限公司,批号K2009336,质量分数≥98%)、原儿茶酸(成都曼思特生物科技有限公司,批号MUST-20110310,质量分数99.78%),甲醇、乙腈均为色谱纯;酒石酸钠钾、七水合硫酸亚铁、磷酸二氢钠、磷酸氢二钠、无水乙醇、甲酸均为分析纯。
-
Ultimate 3000DGLC高效液相色谱仪(赛默飞世尔科技)、超微量分光光度计NP80(德国IMPLEN公司),电子分析天平(上海梅特勒-托利多有限公司),超声波清洗器(上海科导超声仪器有限公司),多功能动态热回流提取浓缩机(上海天巨源设备有限公司),电热恒温鼓风干燥箱(上海跃进医疗器械厂)。
-
取槟榔药材20 kg,粉碎后过3号筛,分别加入10倍、8倍、6倍量的80%乙醇,80 ℃加热回流提取3次,时间分别为2、1.5、1.5 h,合并提取液,过滤,滤液减压回收乙醇,得浸膏。浸膏加入等体积水悬浮,加入到已经处理好的AB-8大孔吸附树脂柱上,纯水洗脱除杂,20%乙醇洗脱,收集20%洗脱液,减压回收乙醇得浸膏,真空60 ℃干燥,制得干燥槟榔多酚提取物1,命名为:BLDF202001。大孔吸附树脂经80%乙醇洗脱,收集洗脱液,减压回收乙醇得浸膏,真空60 ℃干燥,制得干燥槟榔多酚提取物2,命名为:BLDF202002。提取物2加水悬浮,加入到已经处理好的聚酰胺层析柱上,纯水洗脱除杂,80%乙醇洗脱,收集80%洗脱液,减压回收乙醇得浸膏,真空60 ℃干燥,制得干燥槟榔多酚提取物3,命名为:BLDF202003。
-
精密称取没食子酸对照品5 mg,置于5 ml容量瓶中,加蒸馏水超声使其充分溶解,定容,配制成1 mg/ml的对照品溶液。
-
分别精密称取各对照品适量,加甲醇制成含儿茶素1.92 mg/ml、表儿茶素2.46 mg/ml和原儿茶酸1.12 mg/ml的单一对照品储备液。
-
精密称取槟榔多酚提取物10 mg,置于10 ml容量瓶中,加乙醇超声,使其充分溶解,定容,配制成1 mg/ml的供试品溶液。
-
精密称取槟榔提取物50 mg,置于10 ml棕色容量瓶中,用甲醇超声使其充分溶解,定容,配制成浓度为5 mg/ml的供试品储备液。精密吸取供试品储备液2 ml,置于5 ml容量瓶中,用50%的甲醇定容至刻度,摇匀,临用前过0.22 μm微孔滤膜,取续滤液,即得。
-
采用酒石酸亚铁法测定槟榔多酚的含量[14],精密吸取“2.2.1”项下的没食子酸对照品溶液各0.05、0.10、0.15、0.20、0.25、0.30 ml,置于5 ml容量瓶中,加蒸馏水至1 ml,再加入酒石酸亚铁溶液1 ml,用pH 7.5的磷酸缓冲溶液定容,混匀,静置15 min,以蒸馏水为空白参照,在540 nm处测定吸光度值。以吸光度值(A)为纵坐标,浓度(X,μg/ml)为横坐标,绘制标准曲线,得没食子酸对照品的线性回归方程:A=12.44X+0.0731,r=0.999 4,表明没食子酸对照品溶液在9.8~58.8 μg/ml范围内线性关系良好。
-
精密吸取没食子酸对照品溶液0.1 ml,按“2.3.1”项下方法测定吸光度,重复测定6次,没食子酸对照品溶液的吸光度RSD值为0.40%,表明该方法精密度良好。
-
取同一供试品溶液(BLDF202001)6份,按“2.3.1”项下方法测定吸光度,槟榔提取物中总多酚的吸光度RSD值为1.1%,表明该方法重复性良好。
-
取同一供试品(BLDF202001)10 mg,精密称定,按“2.2.3”项下方法制备供试品溶液,分别在0、5、10、15、30 min和1、2、4、8、12、24 h后,按“2.3.1”项下方法测定吸光度,槟榔提取物中总多酚吸光度的RSD值为2.8%,表明该供试品在24 h内稳定性良好。
取同一供试品溶液(BLDF202001)0.1 ml,按“2.3.1”项下方法分别于7、9、11、13、15、17、21 min测定吸光度,槟榔提取物中总多酚吸光度的RSD值为0.93%。表明该供试品溶液在21 min内显色稳定,提示总多酚含量测定应等待7 min之后,在21 min之前完成。
-
取同一批(BLDF202001)已知含量的槟榔多酚提取物,平行6份,按“2.2.3”项下方法制备供试品溶液,取供试品溶液0.1 ml,置于5 ml棕色容量瓶中,加入没食子酸对照品溶液0.1 ml,按“2.3.1”项下方法测定吸光度,没食子酸的加样回收率为98.44%,RSD值为1.4%。。
-
吸取0.3 ml的供试品溶液,按“2.3.1”项下方法测定吸光度,平行测定3次,计算总多酚的含量。3批槟榔提取物中总多酚的含量见表1。
表 1 3批槟榔多酚提取物中总多酚的含量(n=3)
批号 总多酚含量(%) BLDF202001 51.45 BLDF202002 51.82 BLDF202003 52.70 -
色谱柱:AcclaimRSLC120 C18柱(3 mm×150 mm,3 µm);流动相为:乙腈(A)-0.3%甲酸溶液(B),梯度洗脱程序见表2;检测波长:280 nm;流速:1.0 ml/min;柱温:35 ℃;进样量:5 μl。
表 2 梯度洗脱程序
时间(t/min) 流动相A(%) 流动相B(%) 0~5 5~10 95~90 5~7 10~15 90~85 7~9 15~20 85~80 9~12 20~5 80~95 -
分别精密吸取50%的甲醇,混合对照品溶液,供试品溶液各5 μl,按“2.4.1”项下色谱条件进样分析,记录色谱图。结果表明,各峰分离度均较好,各组分的分离度大于1.5,符合定量分析要求,理论板数以儿茶素计,不低于6 000。其中原儿茶酸、儿茶素、表儿茶素的保留时间分别为4.273、7.017和8.433 min,色谱图见图1。
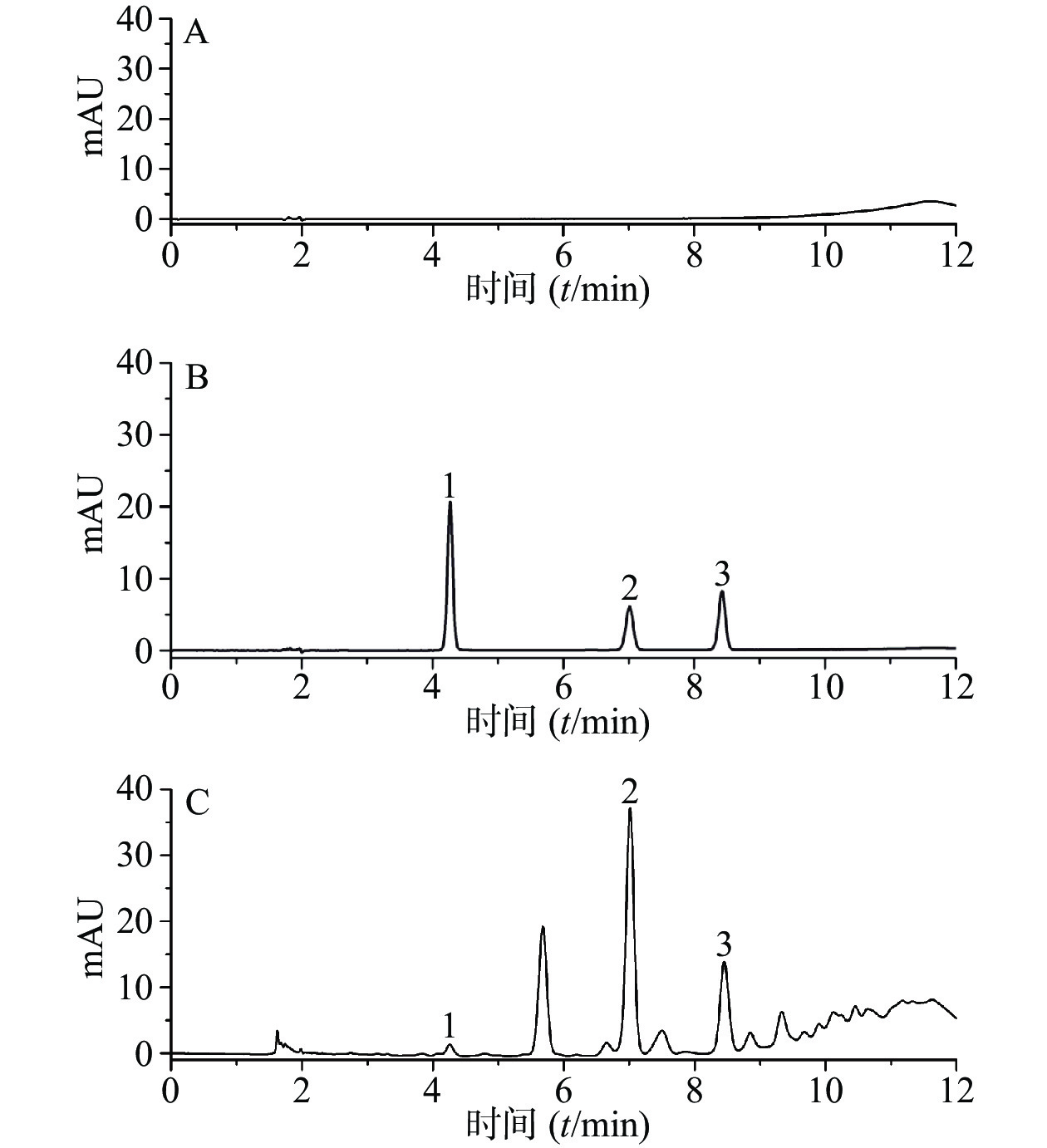
图 1 各成分的HPLC色谱图
-
分别精密吸取“2.2.2”项下儿茶素对照品储备液0.010、0.025、0.05、0.1、0.2、0.4、0.8 ml;表儿茶素储备液0.010、0.025、0.05、0.1、0.2、0.4、0.6 ml;原儿茶酸储备液0.005、0.010、0.025、0.05、0.1、0.2、0.4 ml,用50%的甲醇定容于5 ml容量瓶中,配制成不同浓度的对照品溶液,按“2.4.1”项下的色谱条件进样,以对照品的浓度(X,μg/ml)为横坐标,峰面积(Y)为纵坐标进行线性回归,结果见表3,表明儿茶素、表儿茶素、原儿茶酸分别在相应的线性范围内线性关系良好。
表 3 槟榔多酚提取物中儿茶素、表儿茶素、原儿茶酸的线性方程
成分 回归方程 r 线性范围(μg/ml) 儿茶素 Y=0.0436X−0.0201 0.999 7 3.84~307.2 表儿茶素 Y=0.0415X+0.0395 0.999 8 4.92~295.2 原儿茶酸 Y=0.0833X+0.0113 0.997 7 1.12~89.6 -
取各成分浓度约为20 μg/ml的混合对照品溶液,按“2.4.1”项下的色谱条件连续进样6次,记录峰面积。儿茶素、表儿茶素和原儿茶酸峰面积的RSD值分别为0.65%、0.96%、2.3%,表明仪器精密度良好。
-
取同一供试品溶液(BLDF202001)按“2.4.1”项下的色谱条件分别于0、4、8、12、18 h进样分析,记录峰面积。儿茶素、表儿茶素、原儿茶酸峰面积的RSD值分别为1.1%、1.6%、1.1%,表明供试品溶液在18 h内稳定。
-
取槟榔多酚提取物(BLDF202001)约50 mg,精密称定,平行6份,分别按“2.2.4”项下方法制备供试品溶液,按“2.4.1”项下的色谱条件测定,计算供试品中儿茶素、表儿茶素、原儿茶酸的质量分数分别为5.9%、2.4%、0.10%,RSD值分别为1.0%、2.1%、2.1%,表明该方法的重复性良好。
-
精密称取已知含量的槟榔多酚提取物(BLDF202001)25 mg,置于5 ml容量瓶中,共6份,分别精密加入一定量的儿茶素、表儿茶、原儿茶酸对照品溶液,分别按“2.2.4”项下方法制备供试品溶液,按“2.4.1”项下的色谱条件测定,计算回收率和RSD值。儿茶素、表儿茶素、原儿茶酸的平均回收率为99.17%、101.67%、101.18%;RSD值分别为2.5%、1.2%、1.8%,表明该方法准确度良好。
-
取3批按“2.2.4”项下方法制备的供试品溶液,按“2.4.1”项下的色谱条件分别测定。3个批号的供试品中儿茶素、表儿茶素、原儿茶酸的含量见表4。
表 4 3批槟榔多酚提取物中儿茶素、表儿茶素、原儿茶酸的百分含量(%, n=3)
批号 儿茶素 表儿茶素 原儿茶酸含量 BLDF202001 5.90 2.40 0.10 BLDF202002 0.52 0.43 / BLDF202003 1.30 0.77 /
Determination of total polyphenols and catechins in betel nut polyphenol extract
-
摘要:
目的 建立槟榔多酚提取物中总多酚和儿茶素类含量的测定方法,为槟榔多酚提取物的质量控制提供参考。 方法 采用紫外分光光度法对槟榔提取物中总多酚的含量进行测定;高效液相色谱法对儿茶素类物质进行含量测定。 结果 紫外分光光度法测定槟榔提取物中总多酚含量在9.8~58.8 μg/ml范围内线性关系良好,平均加样回收率为98.44%,RSD为1.4%;在高效液相色谱测定中,儿茶素、表儿茶素和原儿茶酸3种主要成分完全分离,在各自范围内线性关系良好,平均加样回收率为99.17%~101.67%,RSD为1.2%~2.5%。 结论 建立的含量测定方法简单、稳定可靠,可用于槟榔多酚提取物的定量分析,为槟榔多酚提取物的质量控制提供实验依据。 Abstract:Objective To establish a method for the determination of total polyphenols and catechins in betel nut polyphenols extract, and provide reference for the quality control of betel nut polyphenols extract. Methods The content of total phenol in betel nut extract was determined by ultraviolet spectrophotometry. The content of catechins was determined by HPLC. Results The linear range of total polyphenols in betel nuts extract was 9.8~58.8 μg/ml. The three components of catechin, epicatechin and protocatechuic acid were completely separated by HPLC, and the linear relationship was good in their respective ranges, with the recoveries between 99.17% and 101.67%, the RSD between 1.2% and 2.5%. Conclusion The established method is simple, stable and reliable, which could be used for the quantitative analysis of betel polyphenol extract, and provide experimental basis for the quality control of betel polyphenol extract. -
Key words:
- betelnut total polyphenols /
- catechins /
- HPLC /
- UV spectrophotometry
-
药用植物中的次生代谢物是中药药效物质的主要来源,已知的植物次生代谢物生物合成途径有乙酸-丙二酸途径、异戊二烯途径、莽草酸途径等[1],探究植物生物合成的调控因素不仅能提升药材的品质,也为中药有效成分体外合成的工业化提供可能。
植物昼夜节律钟是植物体内应对光照、温度等外界因素随昼夜节律性改变而进化出的一套适应机制[2],对植物生长发育具有不可或缺的作用。大量研究表明,如黄酮类化合物合成的相关结构基因表达,也具有明显的昼夜节律性特点,受昼夜节律钟调控[3-4]。昼夜节律钟的核心部分中央振荡器是MYB蛋白LHY(late elongated hypocotyl)、CCA1(circadian clock associated 1)和伪应答调控蛋白家族(PRRs,pseudo-response regulators)组成,对维持植物昼夜节律的稳定至关重要[5-6]。
PRRs基因都带有2个保守的结构域,氨基端的响应接受结构域(receiver-like domain,RLD),其结构上与磷酸接受域相似,羧基端带有的CCT(Constans/Constans-like/TOC1)结构域,这2个结构域被一个保守程度不高的“可变域”所分隔[7-9]。目前研究发现在CO、CO-like以及TOC1基因中也有带此类结构域,对于植物开花进程有着重要作用[10-11]。
研究表明PRRs家族基因具有增加植物抗逆性,影响植物生物量的积累[12-14],以及调控花发育及衰老等作用[15]。目前对模式植物中PRRs基因的研究较多,如在拟南芥以及水稻中的PRRs基因证明具有调控开花周期的作用[17],但药用植物中PRRs基因的研究则罕见报道。
中药红花(Carthami flos)是菊科植物红花(Carthamus tinctorius L.)的干燥花,有活血化瘀的功效。研究表明,红花主要药效物质为黄酮类化合物,如羟基红花黄色素A(HSYA)、红花素、槲皮素、山奈酚、野黄芩苷[18]等,目前已有对红花黄酮类化合物生物合成的关键基因查尔酮合酶、查尔酮异构酶、糖基转移酶等多种研究[19-20],但调控红花中黄酮类化合物生物合成途径的基因未完全明确。验证昼夜节律钟调控红花中黄酮类化合物的生物合成对提升红花品质意义重大。
本研究依据前期红花转录组数据库基因注释筛选昼夜节律相关基因,并与红花黄酮类化合物的积累量进行相关分析,得到具有调控红花黄酮类化合物生物合成功能的昼夜节律基因,通过qPCR、液质联用(UHPLC-MS)等方法以期阐释PRRs基因的特征与功能,为进一步研究昼夜节律钟调控红花黄酮类化合物的生物合成积累资料。
1. 材料与方法
1.1 材料与仪器
植物材料:云南巍山红花品系(编号ZHH0119),种植于海军军医大学药学院温室,温室条件:恒温25 ℃,昼夜节律为16 h光照/8 h黑暗。仪器与试剂:荧光qPCR仪:ABI7500;Phanta Max Master Mix高保真酶,Trans Top green qPCR super mix,Trans one-step cDNA synthesis super mix逆转录试剂盒(北京全式金公司);Meitler Toledo电子天平(十万分之一量程);高效液相色谱仪:Agilent1290 Infinty LC system; 质谱仪:Agilent 6538 Accurate Mass。
1.2 红花总RNA的提取与cDNA第一链的合成
取用新鲜红花的花冠,根、茎、叶约100 mg研磨成粉。依据Plant Zol说明书提取总RNA,并用紫外分光光度计测定总RNA浓度与质量,样品的A260/A280在1.9~2.1之内可认为符合后续实验要求。将其作为模板逆转录合成cDNA第一链,−20 ℃保存备用。
1.3 红花PRRs基因的筛选与克隆
基于红花转录组数据库,结合基因注释筛选出昼夜节律相关基因,通过红花花冠表达谱获取表达量,与红花代谢组数据库中不同花期黄酮类化合物的积累量进行Pearson分析,获取与黄酮类化合物积累量相关系数r≥0.7的昼夜节律基因进行生物信息学分析,设计引物(表1)进行克隆。PCR产物进行凝胶电泳,回收含有目的条带的凝胶,连接载体后转化大肠杆菌感受态细胞,LBA平板培养,挑取阳性克隆送至上海生工生物工程有限公司测序。
表 1 全长克隆引物引物名称 引物序列 PRR1-F ACCTCAAGGGCCACTGGTTC PRR1-R GTAACAAAAGACTTTCTGAA 1.4 红花PRR1生物信息学分析
在NCBI网站用BLAST在线分析红花PRR1全长序列以及编码蛋白的氨基酸序列进行比对以及同源性分析;在ExPASy在线工具SOMPA得出所编码蛋白的二级结构特征;通过Swiss-Model同源建模预测蛋白三级结构;在ClustalX 2.1软件对其编码蛋白的氨基酸序列与同源蛋白的氨基酸序列进行多重比对分析;使用相邻节点算法(Neighbor-Joining)构建系统进化树,自展分析法(Bootstrap analysis)进行1 000次重复验证进化树可靠性。
1.5 红花PRR1基因表达分布特征分析
以红花根、茎、叶、花4个部位;花冠开花前3 d(Ⅰ期)、开花1 d(Ⅱ期)、开花3 d(Ⅲ期)、开花5 d(Ⅳ期)4个时期[12]以及1 d中8个时间点(6:00、9:00、12:00、15:00、18:00、21:00、0:00、3:00)的花冠第一链cDNA为模板,设计RT-qPCR引物,以Ct60s(KJ634810)作为内参基因,进行qPCR实验,每个样品设3个复孔。
1.6 红花中黄酮类化合物含量测定
取红花Ⅲ期单日内4个时间点(9:00、12:00、15:00、21:00)的花冠烘干至恒重,精密称取5.00 mg置1.5 ml离心管,再加入精密量取的1 ml 60%甲醇,室温放置12 h后超声处理40 min,12 000 r/min,10 min离心取上清液,UHPLC-MS在正、负离子模式下进行检测(表2),参比离子:正离子模式为121.050 9,922.009 8;负离子模式为119.036 3,966.000 7,数据采集与分析使用Agilent MassHunter Analysis4.0软件。以柚皮素、芹菜素、槲皮素、HSYA、山奈酚、红花素、山奈酚-3-O-葡萄糖苷以及野黄芩素为对照品。
表 2 UHPLC-MS分析方法色谱条件 质谱条件 色谱柱:Agilent ZORBAX C18(3.5 μm,2.1 mm ×100 mm) 毛细管电压:3.5 kV 流动相:A:0.1%甲酸-水,B:乙腈 锥孔电压:60 V 流速:0.4 ml/min 离子源温度:350 ℃ 柱温:40 ℃ 碎裂电压:120 V 进样量:0.4 ml 采集范围:50~1 100 m/z 梯度洗脱:0 min,5%B; 4 min,20%B;
6 min, 21% B; 9 min, 26% B; 11 min, 40% B; 15 min, 80% B; 17 min, 95% B1.7 红花PRR1蛋白互作体系研究
1.7.1 红花PRR1蛋白互作网络预测
使用STRING数据库对CtPRR1基因进行蛋白互作网络分析,可信区间设置为0.4。利用Python下的networkX将互作基因进行可视化。
1.7.2 红花酵母双杂cDNA文库构建
将红花cDNA均一化处理得到红花酵母双杂cDNA文库,与载体PGADT7进行同源克隆,产物转化大肠杆菌感受态细胞,涂布LBA平板,37 ℃培养过夜,挑取单克隆菌落进行菌液PCR验证
1.7.3 红花PRR1载体构建与酵母转化
根据PRR1 ORF设计同源引物引入BamHI、XhoI酶切位点。PCR产物经电泳后回收,连接酶切载体pGBKT7转入大肠杆菌,测序验证后完成pGBKT7-PRR1 bait载体的构建。
将pGBKT7-PRR1bait质粒通过PEG/LiAc法转化酵母感受态细胞,涂布SD平板,28 ℃培养,挑取单克隆进行菌液PCR鉴定。将阳性单克隆酵母菌液划线涂布含有X-α-gal和Aba的平板。
1.7.4 酵母双杂文库筛选红花PRR1互作蛋白
将酵母感受态细胞、载体DNA、红花酵母双杂cDNA文库质粒按共转法混合处理后均匀涂布于SD/-His/-Trp/-Ura三缺平板培养,挑取单克隆转移至含有X-α-gal的SD/-His/-Trp/-Ura培养基,扩增阳性质粒用于测序以及酵母双杂验证。
1.8 统计分析
实验结果经SPSS19.0软件处理,计量资料统一表示为(
${{\bar x}} \pm {{s}}$ ),组间比较采用ANOVA分析,以P<0.05为差异具有统计学意义。2. 实验结果
2.1 红花昼夜节律相关基因的筛选
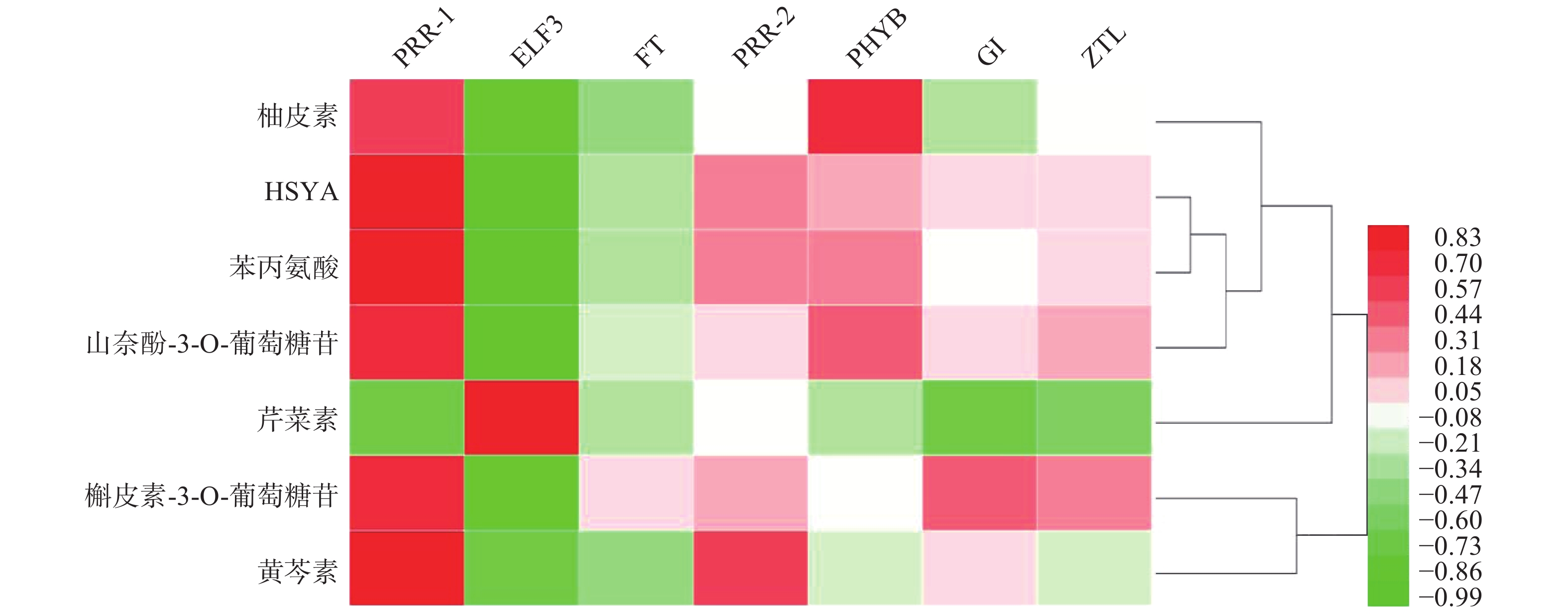
基于红花花冠转录组数据库的基因注释,检索出PRR1、PRR2、ELF3、FT、PHYB、GI、ZTL7个昼夜节律基因,将其在花冠不同时期的表达量与红花代谢组数据库中柚皮素、HYSA、苯丙氨酸、山奈酚-3-O-葡萄糖苷、芹菜素、槲皮苷、野黄芩素、槲皮素-3-葡萄糖苷7个化合物含量进行PEARSON相关性分析,如图1所示,PRR1与红花中主要黄酮类成分的积累量具有良好的相关性(r≥0.7)。
2.2 红花PRR1基因全长克隆与生物信息学分析
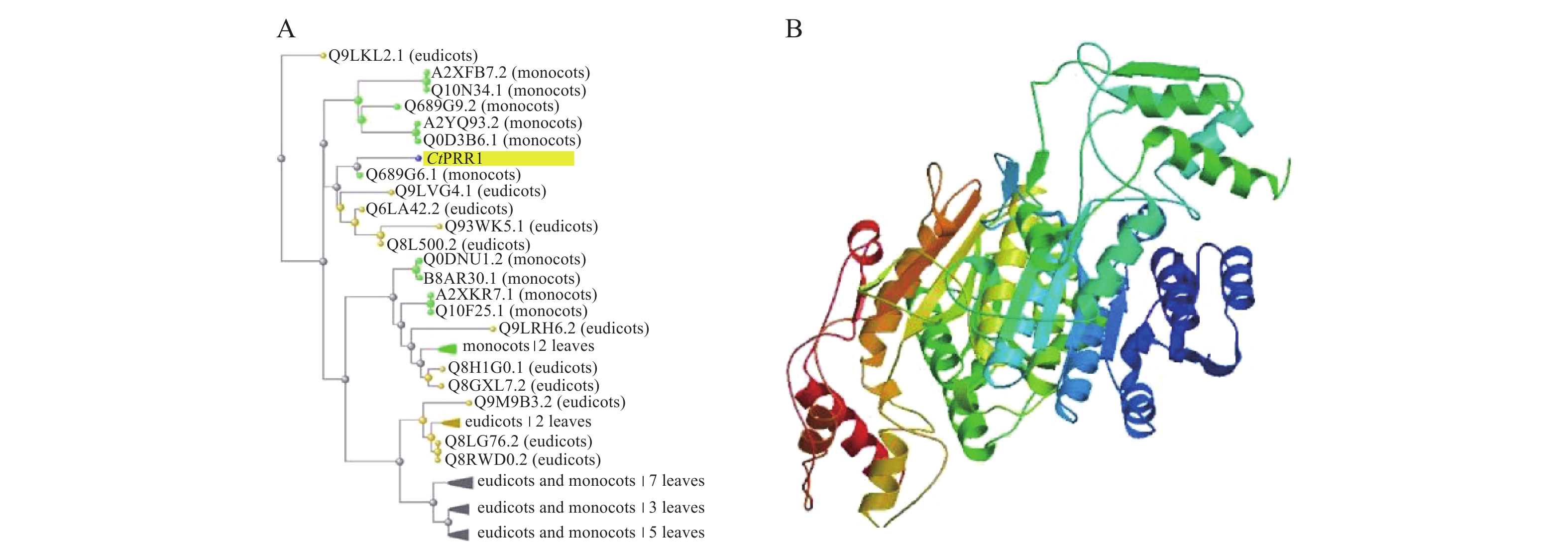
在红花全长转录组数据库中得到的PRR1基因全长序列3 201 bp,ORF FINDER结果显示开放阅读框1 549~2 814 bp,编码421个氨基酸,命名为CtPRR1(GenBank登录号:MW492035),将全长序列进行BLAST,系统发育进化树分析如图2A,CtPRR1氨基酸序列与水稻(Oryza sativa L.)OsPRR73氨基酸序列(A2XFB7.2)同源性最高。Prot-param分析PRR1基因所编码的蛋白质分子式C1900H3039N611O653S15,分子量为45 300,理论pI=8.52,对PRR蛋白质三维结构预测如图2B,Prot Scale分析表明预测PRR1蛋白为亲水性蛋白,无信号肽属非分泌蛋白;蛋白跨膜性分析预测PRR不含有跨膜区域,为非跨膜蛋白。
2.3 CtPRR1基因在红花中的表达特征
qPCR结果表明(图3A)在红花不同部位中,CtPRR1基因在花中的表达量最高,且与根、茎、叶都有显著性差异(P<0.05),对红花花冠Ⅰ期、Ⅱ期、Ⅲ期、Ⅳ期的CtPRR1基因的表达情况进行分析,CtPRR1在Ⅰ期花冠中表达含量最低;Ⅱ期表达水平略有上升,与Ⅰ期无显著性差异;Ⅲ期花冠中表达量明显上升,相比Ⅰ,Ⅱ期具有显著性差异,Ⅳ期花冠中的CtPRR1基因表达水平略微下降(图3B),表明CtPRR1基因在Ⅲ期花冠中转录水平最高,CtPRR1在不同时间表达量的变化与红花黄酮类化合物积累量的变化相符[22]。对红花8个时间点的花冠CtPRR1表达量分析发现CtPRR1表达量在日间6:00至18:00逐渐升高,18:00达峰值,晚间21:00至2:00逐渐下降,3:00最低,表达水平在白天与夜晚两个连续的时间周期内呈现昼夜节律性(图3C)。
 图 3 CtPRR1不同部位及不同时间的相对表达量A.CtPRR1各部位相对表达量; B.CtPRR1不同花期相对表达量; C.CtPRR1红花Ⅲ期单日不同时间点相对表达量。
图 3 CtPRR1不同部位及不同时间的相对表达量A.CtPRR1各部位相对表达量; B.CtPRR1不同花期相对表达量; C.CtPRR1红花Ⅲ期单日不同时间点相对表达量。2.4 红花中黄酮类化合物含量测定
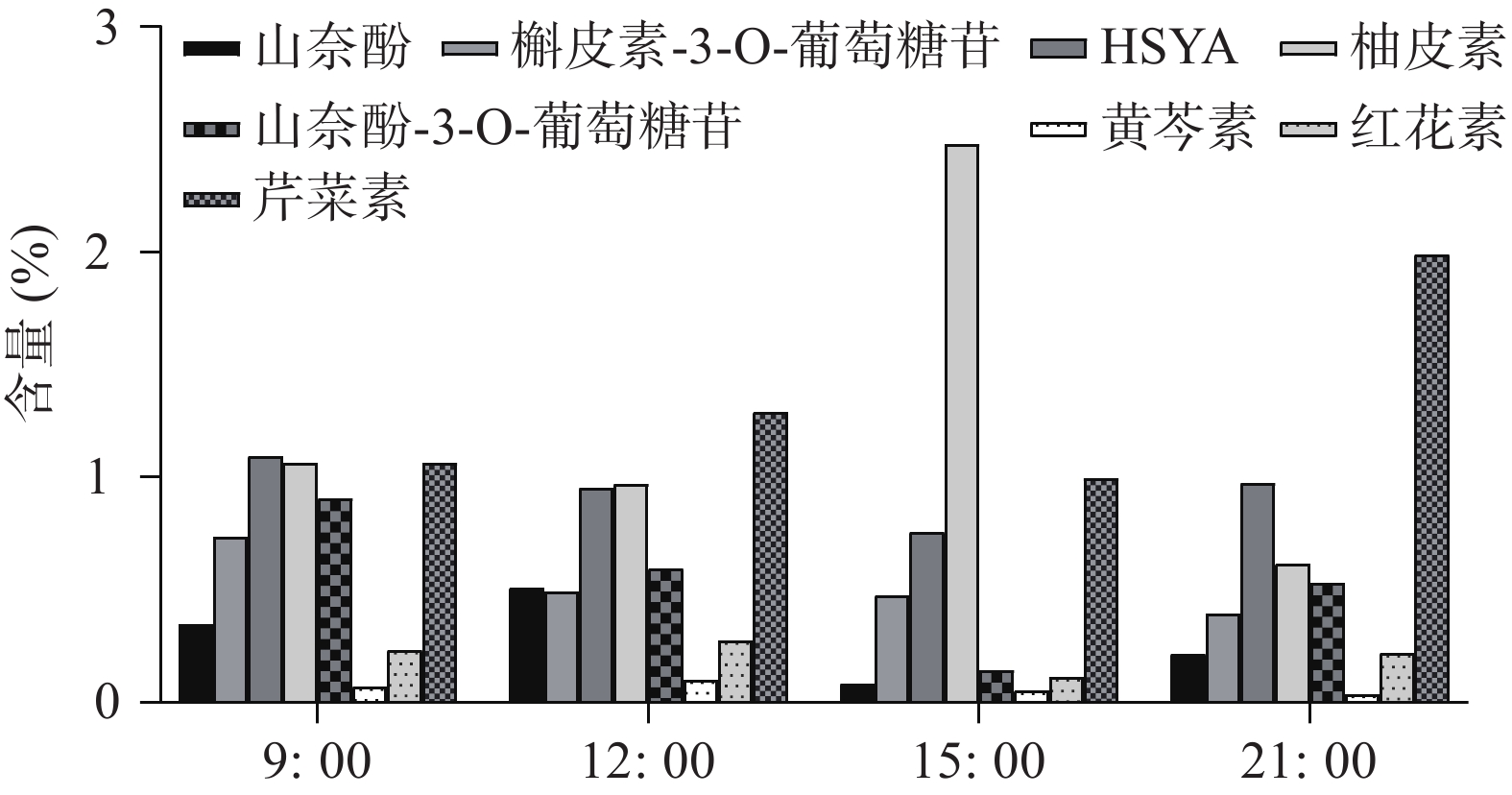
UHPLC-MS检测黄酮类化合物在单日内不同时间的含量变化(图4),除柚皮素外的7个化合物在白天含量逐渐降低,而在晚间含量又升至较高的水平,柚皮素则相反在白天升高,晚间下降,但8个化合物含量变化也呈现昼夜节律性,与同时间点CtPRR1的表达量进行PEARSON分析得到红花素(r=−0.9485)、山奈酚(r=−0.9423)、野黄芩苷(r=−0.9504)、HSYA(r=−0.8372)、山奈酚-3-O-葡萄糖苷(r=−0.8792)、柚皮素(r=0.7415)、芹菜素(r=−0.6652)、槲皮素(r=−0.4876),目前研究认为柚皮素在红花黄酮类化合物生物合成途径的上游[20],而CtPRR1与柚皮素呈正相关,与其他化合物呈负相关,说明CtPRR1对整体黄酮类化合物生物合成具有负调控作用而导致柚皮素的积累量增多,进一步说明CtPRR1参与调控红花黄酮类化合物的合成。
2.5 CtPRR1蛋白互作研究
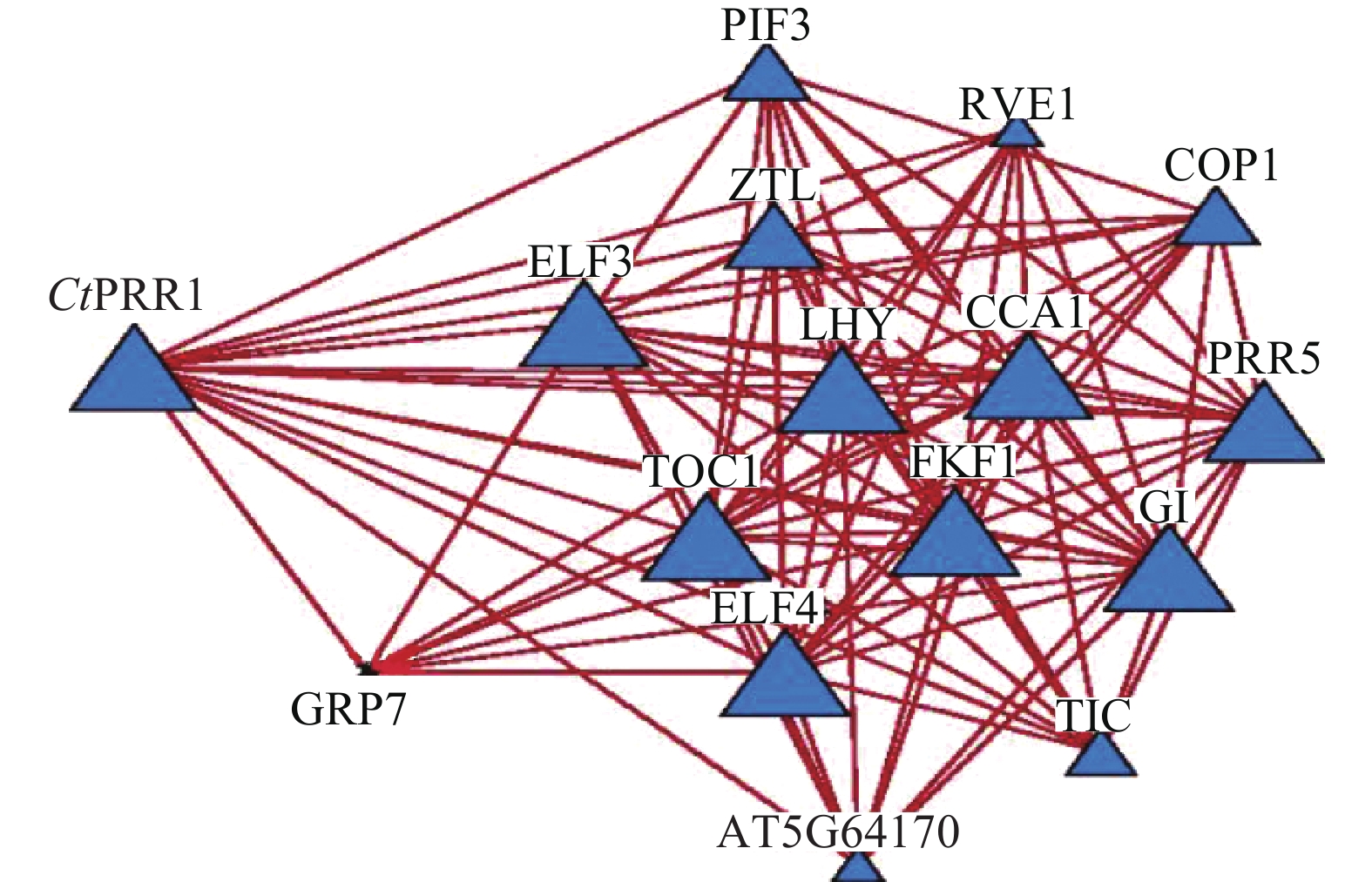
STRING数据库预测红花转录组数据库中CtPRR1的互作蛋白,获得互作蛋白TOC1、PIF3、COP1、ZTL、LHY、ELF4、ELF3、CCA1、GI、PRR5、GRP7、TIC、FKF1、RVE114个,互作关系网络如图5。与PRRs基因共同参与调控植物生长发育,调控植物次生代谢途径。由此获得了较为完善的红花昼夜节律核心元件系统,为进一步探索昼夜节律调控红花次生代谢的分子机制提供了依据。
通过酵母双杂实验发现,转入pGBKT7-PRR bait质粒的酵母细胞在28 ℃生长4 d后情况正常。带有pGBKT7-PRR bait载体的酵母细胞与构建的红花cDNA文库质粒共转后,28 ℃培养2 d后可以观察到蓝色菌斑出现,4 d后出现直径约2 mm的蓝色单克隆菌斑(图6),菌液PCR鉴定为阳性克隆。结果如表3所示,有2个热休克蛋白,3个AP2转录因子,热休克蛋白具有增强植物抗逆性的功能,AP2转录因子广泛参与植物生长发育,调控体内次生代谢。
表 3 酵母双杂筛选互作蛋白序号 蛋白家族名称 预测功能 1 热休克蛋白 广泛参与植物响应外界环境胁迫,生物合成以及生长发育相关的转录调控 2 热休克蛋白 3 AP2转录因子 4 AP2转录因子 5 AP2转录因子 3. 讨论
红花作为常用的活血化瘀中药,对其主要药效物质黄酮类化合物的生物合成途径及调控机制的研究越来越多[20],但目前红花黄酮类化合物生物合成途径及调控机制仍未阐明。
本研究首次克隆了一个PRRs家族基因CtPRR1,生物信息学分析表明其与水稻、拟南芥等其他物种中的PRRs家族基因序列高度相关,说明红花中的PRRs基因具有高度保守性。CtPRR1基因主要在花中表达且开花后第3天时表达量最高,与红花不同花期黄酮类化合物的累积规律一致,存在显著相关。我们认为,CtPRR1调控了红花黄酮类化合物的生物合成。进一步的研究发现,CtPRR1的单日表达量在日间逐渐升高,晚间逐渐下降;随着CtPRR1在单日表达量的升高,芹菜素、槲皮素、HSYA、山奈酚、红花素、山奈酚-3-O-葡萄糖苷以及野黄芩素积累量为白天逐渐降低,晚间逐渐升高。CtPRR1对红花这些黄酮类成分的昼夜节律性积累积起负调节作用;唯CtPRR1与柚皮素的积累量呈正相关,可能与柚皮素处于黄酮生物合成途径的较上游以及参与其他代谢过程并受到其他调控基因的影响有关。
PRRs与CCA1、LHY基因作为昼夜节律系统中的核心元件,对多种植物的昼夜节律钟输出途径中与黄酮合成结构酶基因具有相互作用[22-23],本研究发现红花CtPRR1可能受2个热休克蛋白,3个AP2转录因子的影响对黄酮化合物的积累起负调节作用,丰富了昼夜节律基因调控黄酮类化合物机制的研究资料。
本研究结果为深入研究红花昼夜节律基因对黄酮生物合成途径的调控机制提供了资料。下一步的工作,我们将采用基因过表达以及基因敲除技术结合代谢物分析,进一步验证CtPRR1调控红花黄酮类化合物生物合成的功能及调控网络,为阐明CtPRR1的功能提供重要依据。
-
表 3 槟榔多酚提取物中儿茶素、表儿茶素、原儿茶酸的线性方程
成分 回归方程 r 线性范围(μg/ml) 儿茶素 Y=0.0436X−0.0201 0.999 7 3.84~307.2 表儿茶素 Y=0.0415X+0.0395 0.999 8 4.92~295.2 原儿茶酸 Y=0.0833X+0.0113 0.997 7 1.12~89.6  下载: 导出CSV
下载: 导出CSV
表 4 3批槟榔多酚提取物中儿茶素、表儿茶素、原儿茶酸的百分含量(%, n=3)
批号 儿茶素 表儿茶素 原儿茶酸含量 BLDF202001 5.90 2.40 0.10 BLDF202002 0.52 0.43 / BLDF202003 1.30 0.77 /
下载: 导出CSV
-
[1] 孔丹丹, 李歆悦, 赵祥升, 等. 药食两用槟榔的国内外研究进展[J]. 中国中药杂志, 2021, 46(05):1053-1059. [2] 栾剑, 郭迪, 周晓馥. 槟榔致癌性和毒性的药理学研究进展[J]. 食品与机械, 2019, 35(02):185-189+236. [3] 杨雅蛟, 孔维军, 孙兰, 等. 槟榔化学成分和药理作用及临床应用研究进展[J]. 世界科学技术-中医药现代化, 2019, 21(12):2583-2591. [4] PENG W, LIU Y J, WU N, et al. Areca catechu L. (Arecaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology[J]. Journal of Ethnopharmacology,2015,164:340-356. doi: 10.1016/j.jep.2015.02.010 [5] 易攀, 汤嫣然, 周芳, 等. 槟榔的化学成分和药理活性研究进展[J]. 中草药, 2019, 50(10):2498-2504. doi: 10.7501/j.issn.0253-2670.2019.10.034 [6] FERREIRA I C F R, BAPTISTA P, VILAS-BOAS M, et al. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity[J]. Food chemistry,2007,100(4):1511-1516. doi: 10.1016/j.foodchem.2005.11.043 [7] WANG N, SONG JY, ZHOU G, et al. Mechanism of salidroside relieving the acute hypoxia-induced myocardial injury through the PI3K/Akt pathway[J]. Saudi Journal of Biological Sciences,2020,27(6):1533-1537. doi: 10.1016/j.sjbs.2020.04.035 [8] Lin ES, Li CC. Evaluation of superoxide radical scavenging capacity and reducing power of Areca flower extracts[J]. Journal of Medicinal Plants Research,2010,4:975-981. [9] SARIEF, PRAYOGO GP, LOO YT, et al. Distinct phenolic, alkaloid and antioxidant profile in betel quids from four regions of Indonesia[J]. Scientific reports,2020,10(1):16254. doi: 10.1038/s41598-020-73337-0 [10] 蒋晨凤. 水解对槟榔多酚抗氧化活性的影响及多酚协同抗氧化特性研究[D]. 长沙: 湖南农业大学, 2016. [11] DSOUZA NG, FERNANDES J, D'SOUZA S, et al. In vitro antioxidant activity of leaves extracts of Areca catechu[J]. Research Journal of Pharmacy and Technology,2019,12(4):1536. doi: 10.5958/0974-360X.2019.00254.3 [12] CHAVAN YV, SINGHAL RS. Separation of polyphenols and arecoline from Areca nut (Areca catechu L. ) by solvent extraction, its antioxidant activity, and identification of polyphenols[J]. Journal of the Science of Food and Agriculture,2013,93(10):2580-2589. doi: 10.1002/jsfa.6081 [13] 赵安鹏, 靳婷, 王荣. 槟榔醇提物对H9c2心肌细胞的抗缺氧保护作用[J]. 药学实践杂志, 2019, 37(04):294-298. doi: 10.3969/j.issn.1006-0111.2019.04.002 [14] 宋晶燕. 槟榔多酚的提取分离及其抗高原缺氧药效学的研究[D]. 银川: 宁夏医科大学, 2020. [15] 靳婷. 槟榔提取物抗高原缺氧药效学及其机制研究[D]. 兰州: 兰州大学, 2018. [16] 霍妍, 赵安鹏, 宋晶燕, 等. 槟榔多酚对大鼠高原肺水肿的预防作用[J]. 解放军医学杂志, 2021, 46(10):961-967. doi: 10.11855/j.issn.0577-7402.2021.10.02 [17] GUO Y, PAN Y, ZHANG Z, et al. Study on the browning mechanism of betel nut (Betel catechu L. ) kernel[J]. Food Science & Nutrition,2020,8(4):1818-1827. [18] CHEN W, ZHANG C, HUANAG Y, et al. The inhibiting activity of areca inflorescence extracts on human low density lipoprotein oxidation induced by cupric ion[J]. International Journal of Food Sciences and Nutrition,2012,63(2):236-41. doi: 10.3109/09637486.2011.620946 [19] ADEYANJU A A, ASEJEJE F O, MOLEHIN O R, et al. Protective role of protocatechuic acid in carbon tetrachloride-induced oxidative stress via modulation of proinflammatory cytokines levels in brain and liver of Wistar rats.[J]. Journal of Basic and Clinical Physiology and Pharmacology,2021,3(18):1-12. [20] BERNATONIENE J, KOPUSTINSKIENE D M. The role of catechins in cellular responses to oxidative stress[J]. Molecules,2018,23(4):965. doi: 10.3390/molecules23040965 [21] QU Z, LIU A, LI P, et al. Advances in physiological functions and mechanisms of (−)-epicatechin[J]. Critical reviews in food science and nutrition,2021,61(2):211-233. doi: 10.1080/10408398.2020.1723057 [22] 王明月, 罗金辉, 李建国. HPLC法测定槟榔中的多酚类物质[J]. 天然产物研究与开发, 2011, 23(01):101-104. doi: 10.3969/j.issn.1001-6880.2011.01.024 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4560
- HTML全文浏览量: 1993
- PDF下载量: 24
- 被引次数: 0
