

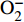
 下载:
下载:

-
聚醚类抗生素是一类重要的微生物次级代谢产物,其结构特征为分子中含有多个环醚单元,且分子一端有羧基,该类抗生素主要由链霉菌产生,具有离子载体性质,容易络合金属离子,常见的聚醚类抗生素有莫能菌素、南昌霉素、尼日利亚菌素等。聚醚类抗生素家族中含有一类特殊的天然产物,即茚满霉素类,该类化合物含有反式四氢茚满环和吡咯酮结构单元,其代表分子是茚满霉素(indanomycin,X-14547A),最早分离自链霉菌Streptomyces sp. NRRL 8167[1]。与其结构相似的化合物还有cafamycin[2]、16-deethylindanomycin[3]及其类似物[4]、homoindanomycin、stawamycin[5-6]和JBIR-11[7]等。茚满霉素类化合物均具有良好的抗菌、杀虫和抗原虫等活性,其特殊的化学结构和显著的生物活性引起了药物学家们的广泛兴趣,不少化学家对其进行全合成,生物学家也对含有该类特殊结构的天然产物的生物合成机制进行研究。
本文就茚满霉素类化合物的天然发现、生物活性、化学合成和生物合成进行总结,为开发该类天然产物的药用价值提供科学基础,为采用组合生物合成的方法对该类化合物进行结构改造提供新思路。
-
在从土壤培养物中寻找新抗生素的过程中,Miller课题组从链霉菌Streptomyces sp. NRRL 8167的发酵液中分离得到了茚满霉素(1),为该家族第一例化合物。化合物1能够将1价和2价阳离子从水溶液中萃取到不相溶的有机溶剂中,另外,与一些只能运输特定单价阳离子(如K+或Na+)的离子载体抗生素不同,化合物1还能通过溶剂屏障(CHCl3)将Rb+和Ca2+从一个水相转移到另一个水相,具有介导跨生物膜转运2价阳离子的特殊能力[8],迄今为止,仅有少数离子载体抗生素(如拉沙里菌素和离子霉素等)具有相似的运输能力[1]。化合物1具有良好的抗菌、杀虫和抗原虫等活性[9-10],具有与其他离子载体抗生素类似的抗菌谱,在体外对G+菌Mycobacterium phlei、Streptomyces cellulosae、Staphylococcus aureus、Bacillus sp. E、Bacillus sp. TA、Sarcina lutea、Bacillus megate- rium、Bacillus subtilis的最低抑菌浓度(MIC)分别为3.1、0.8、0.2、0.2、0.2、0.1、0.1、0.1 μg/ml。研究显示,浓度为100 ppm的化合物1连续使用6 d可以使舞毒蛾和烟草天蛾幼虫的数量减少50%,使玉米穗虫的数量减少33%;浓度为20 ppm的化合物1可以使四龄的埃及伊蚊(Aedes aegypti)的死亡率达100%[1]。王继栋等人发现,化合物1对人乳腺癌细胞(MDA-MB-231)和人肝腺癌细胞(HepG-2)具有一定程度的抑制作用,其IC50值分别为14.01和7.26 μg/ml[11]。
Kliuev等人从产生蒽环类抗生素galtamycin的链霉菌培养液中分离得到了一种新型类似物cafamycin(2)[2],其与化合物1的区别在于2位甲基被乙基取代,18位乙基被脱除。Occolowitz等人从美国蒙大拿州收集的土壤样品中分离出一种新的链霉菌Streptomyces setonii,并从中分离得到了一种新的类似物,即16-deethylindanomycin(3)[3],该化合物对Streptococcus pneumoniae Park I的MIC值为2 μg/ml,对Staphylococcus aureus X1.1、S. aureus V41、S. aureus V400、S. aureus S13E的MIC值为4 μg/ml。当化合物3的浓度为0.31μg/ml时,可以100%抑制柔嫩艾美耳球虫(Emeria tenella)的生长。Zhang等人从海洋链霉菌Streptomyces antibioticus PTZ0016中分离得到了化合物3的类似物,iso-16-deethylindanomycin(4)、16-deethylindanomycin 甲酯(5)和iso-16-deethylindanomycin 甲酯(6),这3种化合物在体外实验中都显示出对金黄色葡萄球菌的抑制活性,其MIC值为4.0~8.0 μg/ml[4]。有趣的是,化合物4和6分别为化合物3和5的C-7手性异构体,这一现象在其他茚满霉素类似物中并不多见,提示该菌株中负责呋喃环形成的酶的立体选择性低。Homoindanomycin (7)分离自菌株Streptomyces galbus,其与化合物1唯一的区别在于2位上的甲基被乙基取代。Miao等人从链霉菌菌株Strepto-myces sp.的液体培养物中分离出一种新的天然产物stawamycin(8),化合物8保留了与化合物1类似的吡咯和四氢茚满结构,但不具有呋喃环,并且双键位置和构型与其他化合物相比也有明显区别,另外,该化合物的绝对构型并没有完全确定。化合物8具有抗人类疱疹病毒EB病毒(epstein-barr virus,EBV)活性,可以抑制病毒转录因子BZLF1与其DNA靶标的结合[5]。Miho等人从绿色链霉菌Streptomyces viridochromogenes的菌丝体中分离得到了JBIR-11(9),是化合物8的衍生物,不同的是化合物9在末端羧基上结合了一分子色氨酸,而且化合物9具有抗肿瘤活性,对人纤维肉瘤HT1080细胞具有生长抑制作用,其IC50值为25 μmol/L[7]。茚满霉素及其天然类似物的结构式如图1所示。
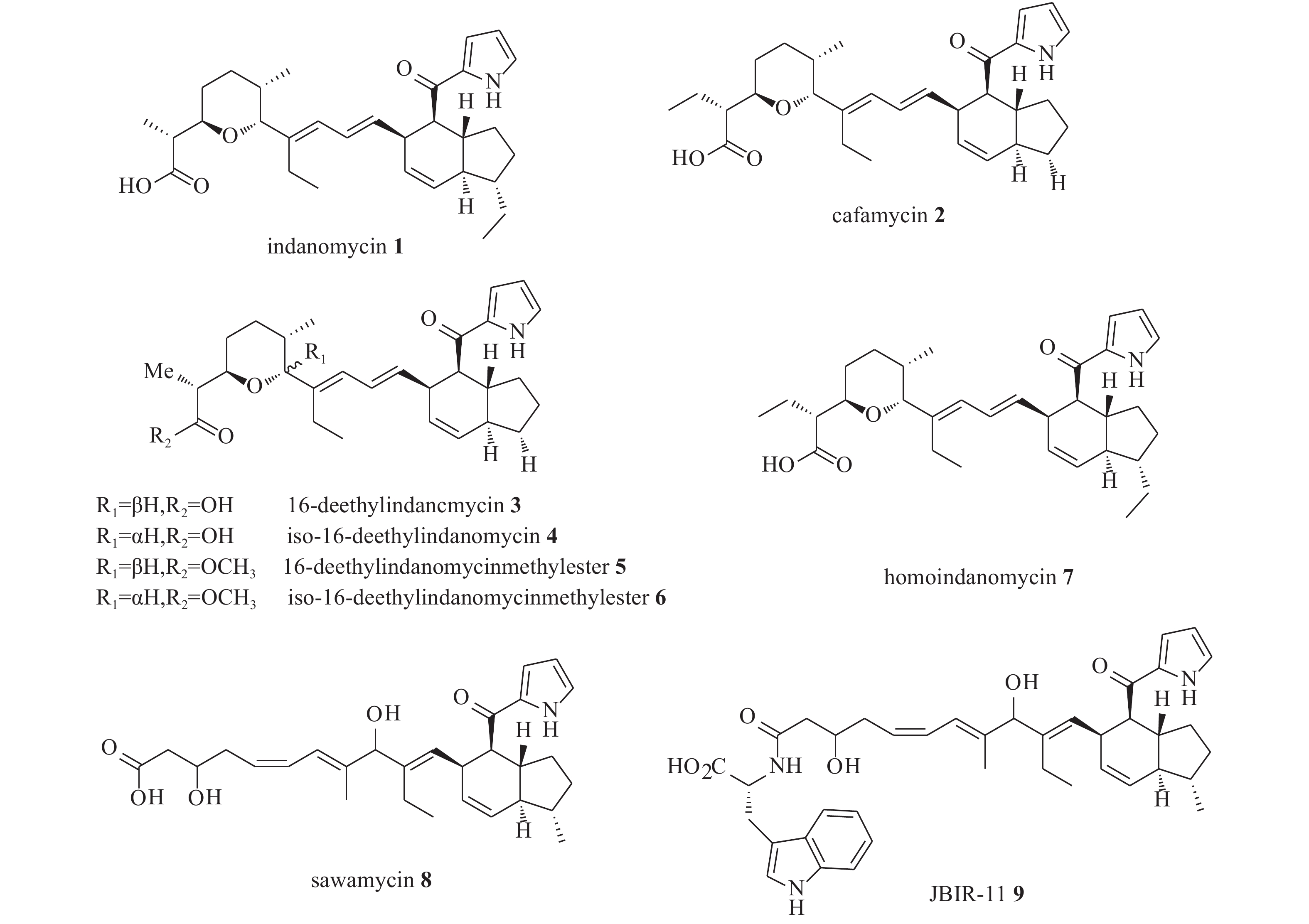
图 1 茚满霉素及其天然类似物
-
由于茚满霉素中不同寻常的结构,如反式四氢茚满环和吡咯酮,其全合成被多次报道[12-15]。Boeckman[16]等人对茚满霉素进行了逆合成分析,选择利用串联Wittig反应与分子内Diels-Alder环加成反应来合成反式四氢茚满环骨架。吡喃醛中间体(10)和磷叶立德(11)发生wittig反应得到中间体(20),随后由分子内[4+2] Diels-Alder环加成反应得到化合物1。
化合物10的构建以活泼醛(12)为原料,在二烷基铜锂的作用下发生羟醛缩合反应得到醇类中间体(13),化合物13在过量臭氧下双键发生断裂后,被硫醚还原得到化合物14。在二叔丁基联苯锂的存在下,化合物14发生锂盐化,最后经PPTS催化得到化合物10(图2)。
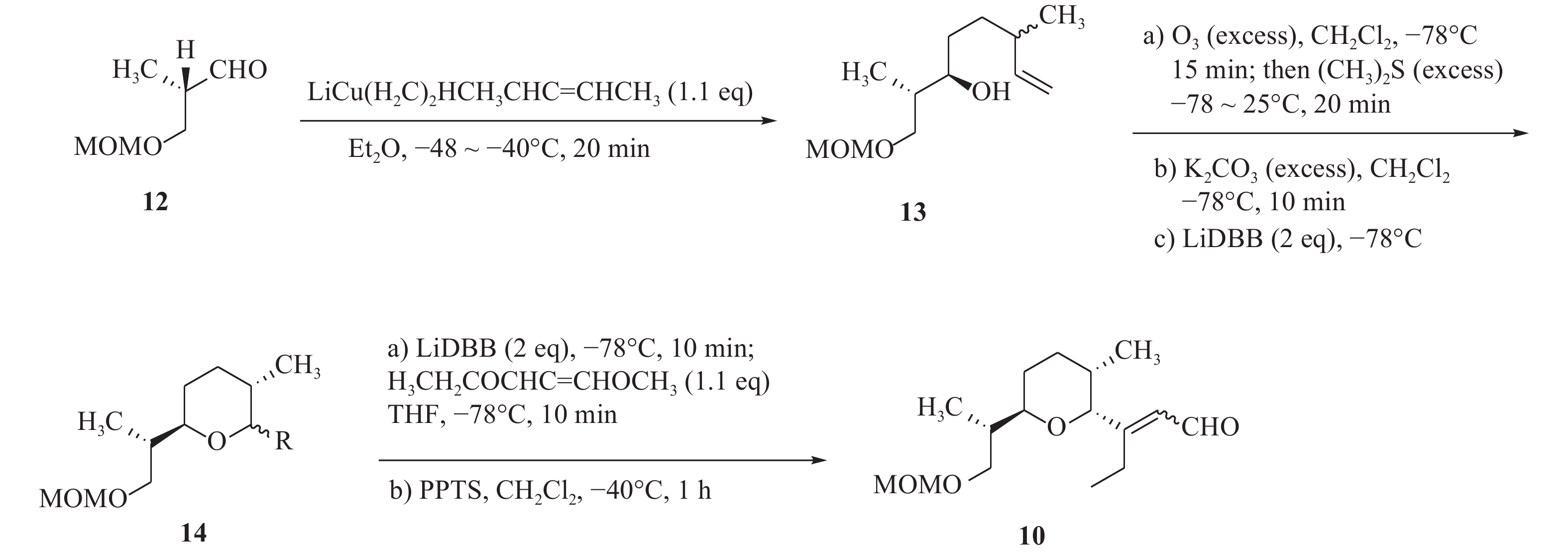
图 2 茚满霉素中间体(10)的化学合成路线
化合物11的合成以二醇化合物(15)为起始原料,在DMAP催化作用下,伯羟基发生硅基烷基化反应,随后与二异丁基氢化铝在−78 ℃发生Claisen重排,得到烯醇(16)。化合物16进行磺基化反应后,以氰化钾作为氰源,合成氰化物(17)。化合物17在四丁基氟化铵和重金属氧化剂PDC作用下,得到α, β不饱和醛(18)。最后将化合物18与格氏试剂乙烯基溴化镁在−78 ℃进行无水无氧反应,由此得到的化合物19再与三苯基膦溴化氢反应,生成化合物11(图3)。
图 3 茚满霉素中间体(11)的化学合成路线
最后,化合物10和11在叔丁醇钾的作用下,发生wittig反应得到中间体20。化合物20被i-Bu2AlH还原为醛,随后在1,2-二氯乙烷中与吡咯酮(21)反应,先后进行wittig反应与分子内Diels-Alder环加成反应,得到化合物22。化合物22在三甲基碘硅烷作用下脱保护,再与过量的三氧化铬发生氧化反应,得到目标化合物1(图4)。
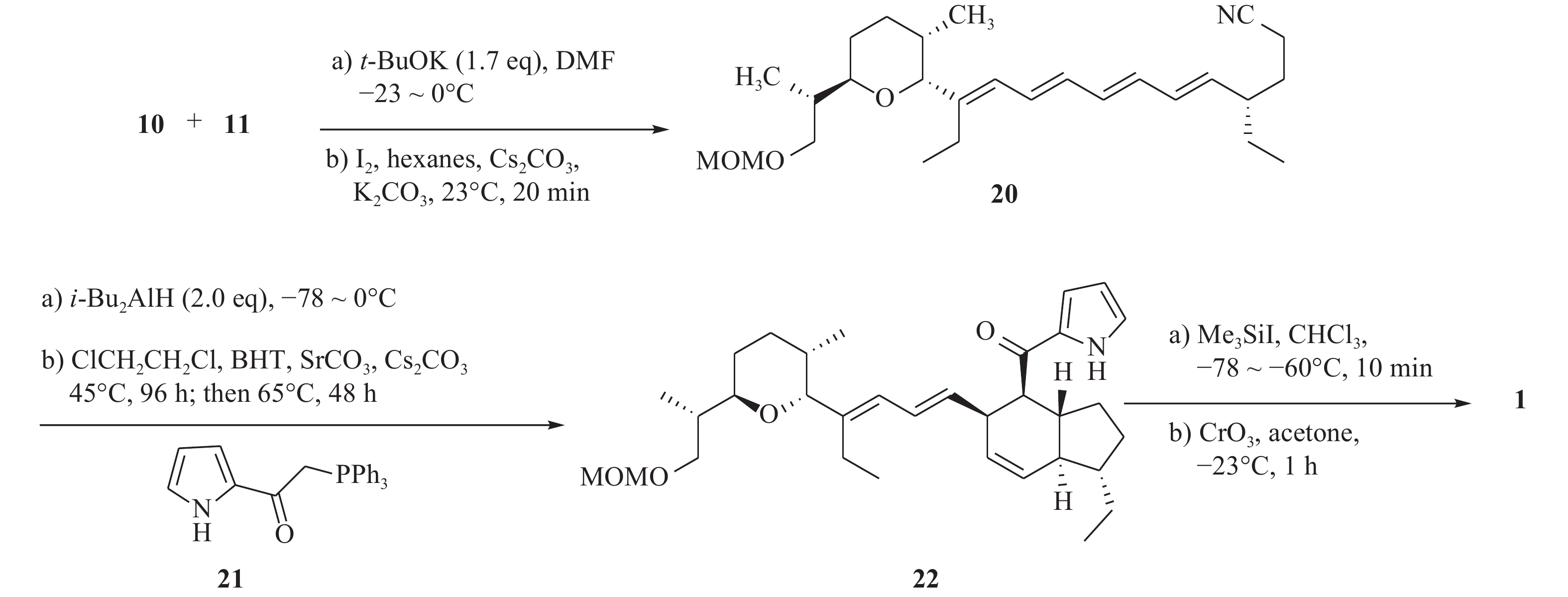
图 4 茚满霉素的化学合成路线
该化合物的全合成历经21步,总体收率不高。反应过程需要使用锂化物、氢化铝、格氏试剂等危险品,还需要–78 ℃的低温无水反应,条件苛刻,并且使用到了剧毒化合物氰化钾、重金属试剂PDC及三氧化铬。因此,该方法对环境不友好,不符合绿色化学的理念,有待进一步的改善。
-
化合物1是杂合了非核糖体肽合成酶-聚酮合酶(NRPS-PKS)装配线的天然产物,Roege等人用13C标记的前体喂养实验确定了其代谢起源,包括1个L-脯氨酸、6个丙二酰辅酶A、2个甲基丙二酰辅酶A和2个乙基丙二酰辅酶A[17-18]。Kelly课题组从抗生素链霉菌NRRL 8167中确认了化合物1的生物合成基因簇idm,其大小为80 kb左右(图5A)[19]。其中16个基因参与了化合物1的生物合成(idm A-P),包括吡咯合成[20](pyrrole biosyn-thesis,idmI-K),调控和抗性基因(idmC,D,G),聚酮合成酶基因(idm L-P),聚酮前体合成基因(idmB,E,F),后修饰基因(idmA,H),另外7个基因经基因敲除实验验证与化合物1的合成无关(orf 1-4,orf 21-23)。起始单元吡咯-2-甲酰CoA是由L-脯氨酸经过脯氨酰依赖的转移酶(idmJ),载体蛋白(idmK)和黄素依赖的L-脯氨酰CoA脱氢酶(idmI)3个酶催化形成的(图5C)。IdmJ基因缺失实验证实idmJ参与了化合物1的生物合成。编码PKS的基因位于idmI-K的下游,包含编码idmL-P 5个酶,根据生物信息分析划分成10个模块:idmL(模块1~3),idmM(模块4和5),idmN(模块6~8),idmO(模块9),idmP(模块10和11)。PKS模块的结构域由酮基合成酶(KS)、酰基转移酶(AT)、脱氢酶(DH)、烯酰还原酶(ER)、酮基还原酶(KR)和酰基载体蛋白(ACP)等结构域组成,根据模块中的结构域分析,推测化合物1的PKS骨架形成与延伸过程如图5B所示。
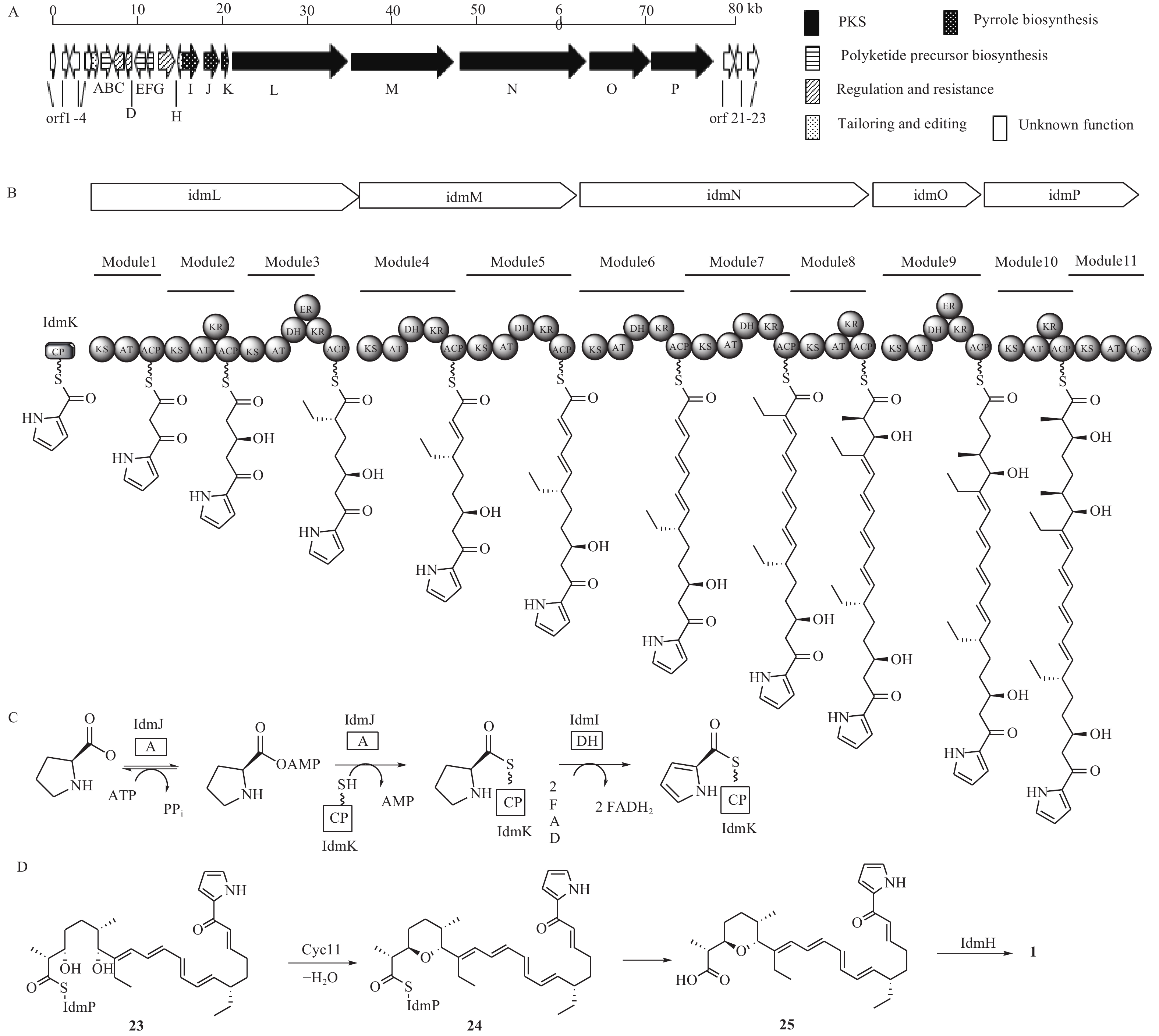
图 5 茚满霉素的生物合成基因簇、PKS模块、前体合成及后修饰过程推测
模块1~10完成了化合物1的PKS骨架搭建,模块11是化合物1生物合成中的特殊PKS模块,它不参与聚酮链的延伸,而且它的模块结构不完整,仅包含一个KS单元、一个AT单元和一个特殊的cyc11单元,其中,cyc11单元与盐霉素(salinomycin)生物合成中的吡喃合成酶(SalBIII)的同源性较高,序列比对显示,cyc11(Asp28和Asp94)含有SalBIII(Asp38和Asp104)活性必需的残基[21-22],因此cyc11可能具有与盐霉素中吡喃形成类似的催化机制[23],即负责催化化合物23的C-3羟基脱水生成α, β-不饱和酮,然后进行C-7羟基的Michael加成产生化合物24。根据生物信息学预测,茚满霉素PKS模块最终对应的产物为含19-OH的中间体,而不是化合物23,因为其PKS模块2中无DH结构域,不能直接产生Δ19(20)双键。但是,该位置上双键需要作为化合物24上的亲双烯体进行[4+2]环加成以产生茚满环,具体哪个酶负责催化形成Δ19(20)双键目前尚不明确。另外,序列比对表明idmH与环化酶SnoaL[24-25]存在很大的相似性,所以负责[4+2]环化形成茚满环结构的酶可能是idmH[19, 26],但目前尚无体外生化反应验证。根据生物信息学推测的茚满霉素后修饰合成路线如图5D所示。
-
含反式四氢茚满环结构的化合物在天然产物中比较少见,自1979年茚满霉素首次报道以来,这类化合物就引起了药物化学家和生物学家的广泛兴趣,本综述总结了其在生物活性、化学全合成以及生物合成等方面获得的研究成果,为该类天然产物的药用价值开发提供科学依据。虽然茚满霉素的生物合成步骤尚未完全阐明,但随着其生物合成基因簇的发现,利用组合生物合成的方法对该类化合物进行结构改造将成为可能。
Research progress on indanomycin natural products
-
摘要: 茚满霉素类天然产物是一类具有反式四氢茚满环(indan)结构的微生物次级代谢产物,该类化合物普遍具有良好的抗菌、杀虫以及抗肿瘤等生物活性,因而引起了药物化学家和生物学家的广泛兴趣。对1979年至今有关茚满霉素类化合物的天然发现、生物活性、化学合成以及生物合成等方面的研究进展进行综述,为该类抗生素的基础和应用研究提供科学参考。Abstract: Indanomycins are a class of secondary metabolites of microorganisms with a trans-tetrahydroindan (indane) ring. These compounds generally have good antibacterial, insecticidal and antitumor biological activities, which have caused wide interest for medicinal chemists and biologists. This review summarizes the research progress of the discovery, biological activity, chemical synthesis and biosynthesis of indanomycin compounds since 1979 and provides scientific reference for the research and development of indanomycin antibiotics.
-
Key words:
- indanomycin /
- indane ring /
- biological activity /
- chemical synthesis /
- biosynthesis
-
1. 电离辐射的危害
随着全球经济的高速发展和科技的不断进步,核工业在军事、医疗等领域得到全面发展,但伴随而来的是对从业人员和附近居民造成严重的辐射危害。
辐射是指能量以电磁波或粒子的形式向外传播的现象,可分为电离辐射和非电离辐射。拥有足够高能量而使原子电离的辐射为电离辐射,它包括X射线、α射线、β射线、γ射线等,具有潜在的致癌性。非电离辐射能量较低,不会电离物质而会使物质内粒子运动,包括红外线、紫外线和微波等[1]。
辐射可引起全身性的放射病,几乎所有系统、器官均可发生病理性改变,其中以神经系统、消化系统和造血器官的改变最为明显,会诱发心血管疾病、糖尿病甚至癌突变。辐射对机体的损伤可分为急性和慢性放射性损伤。短时间内接受高剂量的照射,可引起机体的急性损伤,常见于核事故和放射治疗患者。剂量低于1 Gy时少数会出现轻微症状,剂量在1~10 Gy时,会出现造血型急性放射病;剂量超过10 Gy,会出现高致死率[2]。而长期接受超剂量的全身或局部照射,可引起慢性放射病,如皮肤损伤、造血障碍、白细胞减少、生育功能受损等。此外,辐射还能直接导致视力下降、视网膜脱落,诱发孕妇流产、不育、畸胎、儿童发育不足等[3]。
2 抗辐射天然产物的作用机制[4]
抗辐射药物是指在辐射前或后给予药物预防或治疗,可减轻或修复辐射损伤的药物。现有的抗辐射化学合成药物主要包括细胞因子、含硫化合物和激素类药物[5],因其毒副作用较大而应用受限,近年来天然产物因其毒副作用小、多成分多靶点的独特优势受到广泛的关注。目前认为抗辐射天然产物的作用机制主要有以下4个方面。
2.1 防护DNA损伤
辐射损伤可破坏DNA分子的结构与功能,导致DNA碱基破坏、DNA分子间交联、DNA双链或单链断裂、糖基破坏等。此外,辐射还可导致细胞周期改变以及DNA合成抑制,直接影响细胞增殖。抗辐射天然产物可通过减轻或抑制辐射致细胞周期的缩短,避免或修复DNA损伤而起辐射防护作用。
2.2 清除自由基
人体产生的80%自由基是由水分子组成的。辐射可引起水分子生成强活性的氧化自由基,主要包括·OH、
${\rm{O}}^-_2 $ 、H2O2、·NO等,其中,·OH氧化性最强,可导致组织细胞产生脂质过氧化物[6]。人体由于自由基的产生造成的破坏主要有3个方面:破坏细胞膜;使血清抗蛋白酶失去活性;损伤基因导致细胞变异,如自由基和生物大分子的结合,导致DNA主链断裂或碱基破坏,通过氧化性降解使得多糖链断裂,形成脱氢自由基,破坏细胞膜上的多糖结构[7]。现代研究表明,大多数抗辐射天然产物具有清除多种自由基作用,能降低氧化酶活性,抑制细胞过氧化物的产生。2.3 保护免疫系统
辐射主要损伤骨髓、胸腺和脾脏等免疫器官以及淋巴细胞等。崔玉芳等[8]发现辐射对免疫系统的损伤主要表现为两个特点——早期损伤严重和后期恢复缓慢。在辐射早期脾脏T、B淋巴细胞数量迅速减少,丝裂原反应明显降低,而在受照射1年后,小鼠的免疫组织和外周血淋巴细胞凋亡率与正常水平相比仍较高,小鼠T淋巴细胞免疫功能仍未恢复。促进淋巴细胞增殖,抑制胸腺和脾脏细胞凋亡等是抗辐射损伤的有效途径。
2.4 保护造血系统
造血组织是辐射的敏感组织,机体受到辐射后,造血细胞会出现功能低下甚至死亡现象,其中,造血干细胞、粒系祖细胞、红系祖细胞是辐射攻击的主要靶细胞,外周血细胞的数量随着照射剂量的增加而减少,其形态和功能也会随之发生改变[1]。因此,改善造血微环境,促进白细胞增殖,修复骨髓造血功能等有助于保护造血系统,修复辐射损伤。
3. 抗辐射天然产物
3.1 多糖类化合物
天然多糖包括植物多糖、动物多糖和微生物多糖。它们是一类具有免疫调节、抗肿瘤、抗辐射、抗炎、抗疲劳、抗衰老作用的生物大分子[9]。关于多糖的抗辐射作用的机制尚不清楚,一般认为与多糖的抗氧化,对造血系统的保护,引起免疫系统的效应增强以及诱导产生某些细胞因子等作用有关。
3.1.1 植物多糖
研究表明,大多数植物多糖有较为显著的抗辐射作用,能提高辐射诱导损伤的防护能力,改善辐射诱导的氧化损伤。其辅助保护辐射损伤的作用机制复杂,一般推测与其修复DNA损伤、消除自由基、增强免疫功能等有关[10]。张乃珣等[11]研究发现,酸性黑木耳多糖(AAP)和红松球果多酚的联合使用可以有效地清除体内自由基,降低自由基对体内DNA造成的损伤,显著提高对60Co γ射线诱导氧化损伤的防护能力。此外,白海娜等[12]发现原花青素与黑木耳多糖(AAP-4)同样有协同防护辐射诱导氧化损伤的作用。徐俊杰等[13]研究山药多糖对低强度连续微波辐射致小鼠免疫系统功能损伤的保护作用,发现正常动物组与辐射损伤组相比,不同剂量(200、400、800 mg/kg)的山药多糖可提高巨噬细胞的吞噬指数、T淋巴细胞的增殖刺激指数和血清IgG水平,并降低血清IL-4水平。表明山药多糖能明显改善低强度连续微波辐射对小鼠免疫系统的损害。胡淼等[14]报道,预先给药黑大蒜多糖(150~600 mg/kg)可减轻X射线辐射对小鼠免疫器官和全血白细胞、血小板的影响,提高脾脏的代偿性造血增殖能力,提高抗氧化酶水平,具有较好的辐射防护作用。Zhang等[15]发现大黄多糖(RTP)通过调控Nrf2及其下游蛋白HO-1,显著降低细胞凋亡和炎症因子,从而显著改善辐射诱导的肠道损伤。
3.1.2 动物多糖
国内外学者从动物体内提取出不同种类的多糖,尤其是海洋动物,如虾蟹动物的甲壳质、河蚌多糖、鲍鱼多糖等,具有抗肿瘤、抗病毒、抗氧化、抗辐射等生物活性[16]。
3.1.3 微生物多糖
研究发现微生物中,尤其生活在高压、高辐射环境中的藻类,其多糖有着较为特殊的结构与生理特性,大多有较好的抗辐射效果。Kim等[17]在探讨低分子量岩藻多糖(LMF)对中波紫外线诱导的光老化的保护作用时发现,持续15周的中高剂量(2.0、1.0 mg/cm2)LMF治疗可对受到中波紫外线照射的小鼠光老化起到明显的保护作用,可抑制皱纹形成,皮肤水肿以及中性粒细胞在光老化病灶上的聚集。杨凯业等[18]报道称铁皮石斛多糖、褐藻多糖、灵芝多糖、竹荪多糖在50 mg/L的质量浓度下的复合作用可抑制紫外线辐射诱导的皮肤细胞光老化作用。
3.2 多酚类化合物
植物多酚是广泛存在于植物体内的一类次生代谢产物,包括黄酮类、花色苷类和酚酸类。研究表明,多酚类化合物含有多个酚羟基,具有显著的清除自由基能力,能减轻自由基对机体的伤害,从而起到辐射防护作用[19]。
Lekmine[20]等评价用阿尔及利亚南部特有植物Astragalus gombiformis Pomel地上部分制备的丁醇提取物的药理活性,采用防晒系数(SPF)等评价Astragalus gombiformis Pomel的光保护作用和抗氧化能力,结果表明提取物(SPF=37.78±0.85,SPF值>30的皮肤保护产品被认为是有效的紫外线辐射过滤器)具有良好的紫外线吸收能力,推测主要与其中的黄酮类和酚酸类化合物(主要为水飞蓟素、迷迭香酸、槲皮苷和山柰酚)的紫外吸收能力和抗氧化防御能力有关,具有潜在的辐射防护能力。
3.2.1 黄酮类化合物
黄酮类化合物泛指两个具有酚羟基的苯环(A-与B-环)通过中央三碳原子相互连结而成的一系列化合物,其基本母核为2-苯基色原酮。黄酮类化合物是一类从中草药中提取的天然产物,被认为是一种有效的抗氧化剂,可以调控炎症介质的调节酶或转录因子,通过与DNA的相互作用影响氧化应激,增强基因组稳定,具有神经保护和辐射保护作用[21]。
金银花素(5,7-二羟基黄酮)是从蜂胶、蜂蜜和几种植物中提取的一种黄酮类化合物。Mansour等[22]发现给药金银花素(50 mg/kg)可提高受5 Gy红外线照射雄性Wister大鼠大脑中丙二醛(MDA)水平和半胱氨酸蛋白酶-3(caspase-3)活性,这提示金银花素具有辐射致脑损伤的神经保护作用。Kale等[23]通过组织病理评估,显示槲皮素可显著减少辐射诱导的神经元变性和炎症浸润,揭示了槲皮素对辐射致脑损伤的神经保护作用。
Li等[24]证实芹菜素(4′,5,7-三羟基黄酮)能够一定程度上修复UVB诱导的人表皮角质形成细胞(HEKs)的毛细血管扩张性共济失调的异常突变,从而抑制HEKs细胞凋亡和坏死,表明芹菜素对中波紫外线损伤的HEKs具有新型的保护作用。Prasad等[25]报道水飞蓟宾(silibinin)可以防止中波紫外线诱导的胸腺嘧啶二聚体的形成,通过增加抑癌基因p53水平进而促进DNA修复和(或)启动受损细胞的凋亡。
曲克芦丁(TRX)是一种黄酮类化合物,广泛存在于茶叶、咖啡、谷类食品、各种水果和蔬菜中,具有抗辐射作用,Panat[26]对其清除自由基的能力和抗细胞凋亡活性进行了系统的研究。TRX能清除超氧物、NO和其他模型稳定的自由基,从而保护受辐照的细胞。
有些英国科学家研究发现,每天喝两杯绿茶、吃一个橘子,就可以帮助“电脑族”们抵御计算机辐射[27]。而儿茶素类化合物作为茶叶中的主要功能成分,具有显著的抗辐射作用。茶树中儿茶素类化合物主要包括,儿茶素、表儿茶素、没食子儿茶素、表没食子儿茶素、儿茶素没食子酸酯、表儿茶素没食子酸酯、没食子儿茶素没食子酸酯及表没食子儿茶素没食子酸酯8种单体。其中,表没食子儿茶素没食子酸酯生理活性较为突出,具有抗氧化性和抗细胞凋亡活性,可预防不同刺激对组织的损伤。Korystova等[28]研究发现在对辐射诱导的大鼠主动脉损伤的预防作用中,发现红茶比绿茶更加有效,即使浓度低于1 g/100 ml的红茶也能够有效预防红外线对主动脉造成的损伤。红茶中的儿茶素含量明显低于绿茶,但两种茶中的黄酮醇含量几乎相等。儿茶素、表没食子儿茶素和表没食子儿茶素没食子酸酯可增加大鼠主动脉的氧化应激,而黄酮醇可降低辐射诱导的氧化应激。因此,红茶药效的提高是由于儿茶素含量的降低使黄酮醇的正向调节作用更大程度地得到发挥所致。
3.2.2 酚酸类化合物
酚酸类化合物系指具有多羟基的芳香羧酸类化合物,主要以糖、酯以及有机酸的形式存在于植物中,现代研究表明酚酸类化合物能够清除体内多种自由基,具有良好的抗氧化活性和潜在的辐射防护作用。
Milton等[29]报道,鱼腥草细胞培养物的甲醇提取物因细胞产生酚类次生代谢物而具有潜在的光保护作用,结果显示鱼腥草细胞的甲醇提取物(310~2500 g/ml)能够显著提高受紫外线照射的3T3-Swiss白化成纤维细胞活力。提取物的LC-MS化学分析表明,其总酚和总酚酸含量(主要为没食子酸和毛蕊花苷)较高,具有特征的紫外吸收峰(第一和第二波段的峰值分别为294和330 nm),能够抵消紫外线对皮肤的有害影响。
Abozaid等[30]报道肉桂酸纳米颗粒可作为一种辐射诱导胰腺炎的氧化还原信号通路的调节剂,首先用I-精氨酸和γ射线诱导大鼠患急性胰腺炎,口服肉桂酸纳米颗粒(CA-NPs)后,急性胰腺炎的严重程度及血清淀粉酶和脂肪酶水平均降低。同时,胰腺组织的MDA水平显著降低,谷胱甘肽的消耗显著恢复,caspase-3水平降低,可明显改善胰腺组织损伤或凋亡。因此,肉桂酸纳米颗粒对辐射诱导的急性胰腺炎具有较好的治疗潜力。Liu等[31]研究发现姜黄素(Cur)对长波紫外线辐射诱导的人皮肤成纤维细胞(HDFs)光老化具有一定的保护作用。Zhang等[32]发现白藜芦醇通过激活Sirtuin1 (Sirt1,组蛋白去乙酰化酶家族成员之一,可减轻炎症损伤)减轻辐射诱导的小鼠肠道损伤。周瑞芳等[33]研究表明,丹酚酸B可减轻γ射线辐射诱导的造血系统损伤和骨髓细胞的DNA及蛋白质的减少,恢复小鼠免疫系统的辐射损伤,具有显著的抗γ射线辐射作用。
3.2.3 花色苷
花色苷是花青素和糖以糖苷键结合而成的一种化合物,广泛存在于植物的花、果实、茎、叶和根器官的细胞液中,起到保护植物抗氧化的作用。其抗氧化和消除自由基能力可防护不同射线辐射,能够发挥独特的生理效应。
Fernandes等[34]发现花色苷家族成员(矢车菊色苷、锦葵色苷及其衍生色素)具有促进皮肤维持健康的活性,研究表明大部分化合物能够抑制金黄色葡萄球菌和铜绿假单孢菌菌株的生长繁殖,减少HEKs和HDF活性氧的产生,抑制皮肤降解酶的活性且无细胞毒性作用,具有一定的紫外线过滤作用。
Targhi等[35]研究黑桑花色苷对大鼠肝组织和骨髓细胞的辐射防护作用,以 60Co γ射线远距放射(3 Gy和6 Gy)建立大鼠辐射损伤模型,随后腹腔注射200 mg/kg的黑桑花色苷,结果显示黑桑花色苷可降低大鼠肝脏MDA和SOD的水平,降低γ射线照射对大鼠骨髓细胞和肝脏的遗传毒性和细胞毒性,有潜在的辐射保护作用。
3.3 皂苷类化合物
皂苷(saponin)类化合物是苷元为三萜或螺旋甾烷类化合物的一类糖苷,存在于人参、桔梗、刺五加等许多中草药中,在增强免疫、抗肿瘤、抗炎等方面具有显著的生物活性。研究表明人参皂苷的抗辐射机制与清除自由基、抗氧化活性,与其对心血管系统、免疫系统的保护作用以及对细胞凋亡的抑制作用有关[36]。
Wen[37]等研究黄芪甲苷对中波紫外线诱导的大鼠真皮成纤维细胞早衰的抗光老化作用,结果显示黄芪甲苷不仅能通过激活细胞外调解蛋白激酶ERK和丝裂原活化蛋白激酶p38信号抑制中波紫外线诱导的胶原-I的降解,还通过激活细胞自噬增加胶原-I的积累,从而保护中波紫外线诱导的光老化细胞,表明黄芪甲苷在抗光老化治疗中的潜在优势。
Wang等[38]分析柴胡皂苷-d (SSd)对肝癌细胞自噬活性和放射敏感性的影响,SSd通过抑制mTOR磷酸化促进肝癌细胞自噬,增加辐射诱导的肝癌细胞凋亡并且抑制肝癌细胞的增殖,为肝癌的放射增敏治疗提供了一种可能的途径。
Kim等[39]研究知母皂苷A-III(TA-III)对中波紫外线诱导的HEKs和HDF侵袭效应的保护作用时发现,TA-III在非细胞毒性剂量下(50 nmol/L)以剂量依赖的方式抑制中波紫外线诱导的环氧合酶-2(COX-2)、基质金属蛋白酶-9(MMP-9)转录和蛋白表达水平,降低中波紫外线诱导的原代皮肤细胞的侵袭,组织肿瘤坏死因子-α(TNF-α)、白介素-6(IL-6)和COX-2在HEKs中的过度表达,表明其具有光保护剂的开发潜力。
3.4 其他
除了上述多糖类、多酚类以及皂苷类化合物,天然产物中的许多其他化合物同样具有良好的辐射防护作用,包括维生素类、蛋白类、无机成分、稀有元素等。
Rostami等[40]研究发现预先摄入硒和维生素E能够对X射线辐射引起的遗传损害起到一定的防护作用。段一凡等[41]报道茶叶籽不饱和脂肪酸对中波紫外线诱导的HEKs损伤具有保护作用。Jaisin等[42]研究发现胡椒碱(10~40 µmol/L)预处理可抑制中波紫外线诱导的炎症信号通路,减弱HEKs的细胞毒性并且抑制其凋亡。这提示胡椒碱的抗炎作用能保护HEKs免受中波紫外线辐射的损伤,可作为一种紫外线辐射诱导皮肤炎症的有效治疗手段。
4. 结语
近年来,国内外越来越重视辐射损伤的防护,抗辐射药物的寻找也变得十分紧迫。而与传统的化学合成药物相比,天然来源的药物具有活性高、选择性强、毒副作用小等优点,作为抗辐射药物有着广阔的开发前景。但是抗辐射天然产物的筛选方法耗时耗力,因此建立高通量、高专属性的抗辐射天然产物筛选方法意义重大。此外,对已有的天然产物进行结构改造,以期获得抗辐射活性更高或毒副作用更小的衍生物以及提高抗辐射天然产物的提取纯化效率等皆是未来抗辐射天然产物研究的重点和难点。
-
[1] LIU C M, HERMANN T E, LIU M, et al. X-14547A, a new ionophorous antibiotic produced by Streptomyces antibioticus NRRL 8167. Discovery, fermentation, biological properties and taxonomy of the producing culture[J]. J Antibiot,1979,32(2):95-99. doi: 10.7164/antibiotics.32.95 [2] MURENETS N V, KUDINOVA M K, KOROBKOVA T P, et al. Kafamycin: a new pyrrol ether antibiotic[J]. Antibiot Med Biotechnol,1987,32(11):811-814. [3] LARSEN S H, BOECK L D, MERTZ F P, et al. 16-Deethylindanomycin (A83094A), a novel pyrrole-ether antibiotic produced by a strain of Streptomyces setonii. Taxonomy, fermentation, isolation and characterization[J]. J Antibiot,1988,41(9):1170-1177. doi: 10.7164/antibiotics.41.1170 [4] LIAN X Y, ZHANG Z Z. Indanomycin-related antibiotics from marine Streptomyces antibioticus PTZ0016[J]. Nat Prod Res,2013,27(23):2161-2167. doi: 10.1080/14786419.2013.793688 [5] MIAO S. Stawamycin, A new pyrroloketoindane natural product from the cultures of Streptomyces sp[J]. Tetrahedron Lett,1995,36(32):5699-5702. doi: 10.1016/00404-0399(50)11274- [6] DIAS L C, JARDIM L S A, FERREIRA A A, et al. Towards the total synthesis of Stawamycin. Synthesis of C11-C21 fragment[J]. J Braz Chem Soc,2001,12(4):463-466. doi: 10.1590/S0103-50532001000400003 [7] IZUMIKAWA M, KOMAKI H, HASHIMOTO J, et al. Stawamycin analog, JBIR-11 from Streptomyces viridochromogenes subsp. sulfomycini NBRC 13830[J]. J Antibiot,2008,61(5):326-329. doi: 10.1038/ja.2008.47 [8] FAYE D, MBAYE M D, COUFOURIER S, et al. Zinc mediated straightforward access to diacylpyrroles[J]. Comptes Rendus Chimie,2017,20(5):492-499. doi: 10.1016/j.crci.2017.01.003 [9] GUMILA C, ANCELIN M L, JEMINET G, et al. Differential in vitro activities of ionophore compounds against Plasmodium falciparum and mammalian cells[J]. Antimicrob Agents Chemother,1996,40(3):602-608. doi: 10.1128/AAC.40.3.602 [10] ZHANG D, NAIR M G, MURRY M, et al. Insecticidal activity of indanomycin[J]. J Antibiot,1997,50(7):617-620. doi: 10.7164/antibiotics.50.617 [11] 张鑫, 姜南, 沈雪莉, 等. 土壤链霉菌HS-HY-197抗肿瘤代谢产物的研究[C]//2010年中国药学大会暨第十届中国药师周论文集. 天津, 2010: 4394-4398. [12] AZZI N, GRIFFEN E, LIGHT M, et al. An enantioselective desymmetrisation approach to C9-substituted trans-hydrindene rings based on a diastereotopic group-selective intramolecular Diels-Alder reaction[J]. Chem Commun (Camb),2006(47):4909-4911. doi: 10.1039/B607488J [13] WHITNEY R A. Cyclic hydroboration of geraniol derivatives: a synthesis of the left-hand portion of X-14547A[J]. Can J Chem,1986,64(4):803-807. doi: 10.1139/v86-132 [14] CLARKE S L, MCSWEENEY C M, MCGLACKEN G P. Investigation of a novel diamine based chiral auxiliary in the asymmetric alkylation of ketones[J]. Tetrahedron: Asymmetry,2014,25(4):356-361. doi: 10.1016/j.tetasy.2014.01.006 [15] ROUSH W R, MYERS A G. Antibiotic X-14547A: total synthesis of the right-hand half[J]. J Org Chem,1981,46(7):1509-1511. doi: 10.1021/jo00320a060 [16] BOECKMAN R K, ENHOLM E J, DEMKO D M, et al. An efficient enantioselective total synthesis of (-)-X-14547A (indanomycin)[J]. J Org Chem,1986,51(24):4743-4745. doi: 10.1021/jo00374a054 [17] ROEGE K E, KELLY W L. Biosynthetic origins of the ionophore antibiotic indanomycin[J]. Org Lett,2009,11(2):297-300. doi: 10.1021/ol802422n [18] RAVINDRAN A, SUNDERRAJAN S, PENNATHUR G. Phylogenetic studies on the prodigiosin biosynthetic operon[J]. Curr Microbiol,2019,76(5):597-606. doi: 10.1007/s00284-019-01665-0 [19] LI C X, ROEGE K E, KELLY W L. Analysis of the indanomycin biosynthetic gene cluster from Streptomyces antibioticus NRRL 8167[J]. ChemBioChem,2009,10(6):1064-1072. doi: 10.1002/cbic.200800822 [20] NIEHS S P, DOSE B, SCHERLACH K, et al. Genome mining reveals endopyrroles from a nonribosomal peptide assembly line triggered in fungal-bacterial symbiosis[J]. ACS Chem Biol,2019,14(8):1811-1818. doi: 10.1021/acschembio.9b00406 [21] MIYANAGA A. Michael additions in polyketide biosyn-thesis[J]. Nat Prod Rep,2019,36(3):531-547. doi: 10.1039/C8NP00071A [22] LUHAVAYA H, DIAS M V B, WILLIAMS S R, et al. Enzymology of pyran Ring A formation in salinomycin biosynthesis[J]. Angew Chem Int Ed,2015,54(46):13622-13625. doi: 10.1002/anie.201507090 [23] KNIRSCHOVÁ R, NOVÁKOVÁ R, FECKOVÁ L, et al. Multiple regulatory genes in the salinomycin biosynthetic gene cluster of Streptomyces albus CCM 4719[J]. Folia Microbiol (Praha),2007,52(4):359-365. doi: 10.1007/BF02932090 [24] SULTANA A, KALLIO P, JANSSON A, et al. Structure of the polyketide cyclase SnoaL reveals a novel mechanism for enzymatic aldol condensation[J]. EMBO J,2004,23(9):1911-1921. doi: 10.1038/sj.emboj.7600201 [25] KLYMYSHIN D A, STEFANYSHYN O N, FEDORENKO V A. Role of genes snoaM, snoaL, and snoaE in the biosynthesis of nogalamycin in Streptomyces nogalater Lv65[J]. Cytol Genet,2015,49(3):152-157. doi: 10.3103/S0095452715030081 [26] ROMMEL K R, LI C X, KELLY W L. Identification of a tetraene-containing product of the indanomycin biosynthetic pathway[J]. Org Lett,2011,13(10):2536-2539. doi: 10.1021/ol200570u -

 点击查看大图
点击查看大图
计量
- 文章访问数: 10925
- HTML全文浏览量: 2059
- PDF下载量: 75
- 被引次数: 0
