

 下载:
下载:


 下载:
下载:


-
阿霉素(adriamycin,ADR)是蒽环类DNA拓扑异构酶Ⅱ抑制剂,疗效确切,抗肿瘤谱广,临床上常用于治疗血液系统肿瘤和胃癌、肝癌等多种实体瘤。但阿霉素也有多种严重的不良反应,其心脏毒性常导致患者充血性心力衰竭,故临床应用受到很大的限制。在使用阿霉素治疗肿瘤的同时,给予具有心脏功能保护作用的药物对抗其副作用是一种有效的办法[1]。因此,寻找抗阿霉素心脏毒性损伤的药物已成为近年来的研究热点。参附汤出自明代医家薛己《正体类药》,由人参和炮附子按3∶2配伍组成,具有益气回阳固脱的功效,是传统中医治疗慢性心力衰竭的经典名方。参附汤的药理作用机制尚不明确,阿霉素心脏毒性的发生机制也还在探索中。研究表明参附汤可通过调节microRNAs的表达,抑制线粒体途径的细胞凋亡,改善血流动力学相关指标,抑制心肌细胞自噬等途径发挥心脏保护作用[2-4]。这些研究对参附汤的药理学作用机制进行了有益的探索,但仍不够深入,目前,鲜见从代谢组学角度对参附汤的药理作用机制进行整体研究的相关报道。
代谢组学是一种快速发展的系统生物学方法,能够对暴露于致病因素或药物治疗的复杂生物系统的海量代谢产物提供整体的代谢状态分析,正好契合中医“多成分、多靶点、多途径”整合调节的作用特点,是研究中药的药效成分和药理作用机制的强有力手段[5-6]。因此,本研究采用GC-MS血清代谢组学技术,检测经参附汤治疗的阿霉素心肌病小鼠血清中代谢物的变化,通过多变量统计分析寻找潜在生物标志物,综合评价参附汤抗阿霉素心肌病的作用,以期从整体、多靶点的层面,深入阐释参附汤抗阿霉素心肌病的作用机制。
-
Thermo-Finnigan Trace DSQ气相色谱-质谱联用仪(Thermo Electron Corporation);Vevo-3100LT高分辨超声显像系统(加拿大Visual Sonics公司)。
-
乳酸脱氢酶(LDH)、肌酸磷化酶-同功酶MB试剂盒(CK-MB)(南京建成生物科技有限公司);三甲基氯硅烷(TMCS)、甲氧胺盐酸盐、N-甲基-N-三甲基硅烷基三氟乙酰胺(MSTFA)、甲醇、吡啶、正庚烷、柠檬酸(美国Sigma-Aldrich公司);酪氨酸、苯丙氨酸、丙氨酸、油酸、硬脂酸、乳酸、琥珀酸、缬氨酸、甘油-3-磷酸、苹果酸、花生四烯酸(上海阿拉丁试剂有限公司);炮附子(产地:四川)、人参(产地:吉林)购自西安市北京同仁堂药房。
-
BALB/c雄性小鼠24只,体重(20±3)g,空军军医大学实验动物中心提供。小鼠饲养条件:恒温(25±1)℃,12 h/12 h昼夜的房间,自由进食标准饲料和纯净水。
-
称取人参60 g,炮附子40 g,采用6倍体积的沸水法提取2次,每次提取2 h,合并2次提取液,减压浓缩至浓度为0.45 g/ml(相当于含人参0.3 g和制附子0.2 g)的参附汤溶液。
-
经过1周的适应性饲养后,24只BALB/c小鼠随机分为3组(正常组、阿霉素模型组、参附汤治疗组),每组8只。模型组:在第1、3、5、7、9、11 d分别腹腔注射阿霉素2、2、3、3、3、3 mg/(kg·d),累计剂量16 mg/kg。参附汤治疗组:在阿霉素造模前3 d,每天灌胃给予参附汤原液1 g/(kg·d),造模的同时继续给药,造模后再连续给药12 d。正常组:腹腔注射等量的生理盐水。
-
各组最后一次给药后72 h,将小鼠用1.5%~2%的异氟烷麻醉,然后行超声心动图检查,评估左心室收缩功能,射血分数(EF)和缩短分数(FS)通过标准公式计算而得。
-
小鼠眼眶取血,血样在室温下放置45 min,4℃下3 000 r/min离心15 min,所得血清一部分用于LDH、CK-MB酶分析,另一部分置于−80 ℃冰箱冻存,用于代谢组学分析。代谢组学质量控制样品(QCs)取自合并等量不同小鼠的血清。
-
将冻存的血清于室温下融化,取100 μl血清,加入300 μl甲醇,涡旋1 min,在冰水浴中孵育10 min,4℃下14 000 r/min离心15min,放置于4℃冰箱中保存。GC-MS分析前,取上清液转移到玻璃瓶,室温下氮气吹干。衍生化:在血清干燥物中加入15 mg/ml的甲氧胺吡啶溶液50 μl,涡旋5 min,70℃ 烘箱放置1 h。加入50 μl MSTFA(含有1% TMCS),涡旋5 min,室温放置1 h;加入150 μl正庚烷,涡旋30 s,4 000 r/min离心10 min,取上清液150 μl至样品瓶。
-
GC-MS分析采用TR-5MS毛细管柱(30 m×250 μm,0.25 μm),进样量1 μl,分流比为10:1,载气为氦气,柱流速为1 ml/min,进样口温度为260 ℃;接口温度为260 ℃;离子源温度为200 ℃;四级杆温度150℃,EI离子源70 eV;采用全扫描模式,m/z:60~600;程序升温条件:70℃保持3 min,以4℃/min升至220 ℃,然后以12℃/min升至310℃后,保持10 min。样品分为8个小组进样,每小组的3个样品分别取自正常组、阿霉素组、参附汤治疗组,每个小组内采用随机方式进样,每个小组之间进样一次QCs,用于监测分析系统的稳定性。
-
原始数据经ThermoXconvert软件转为NetCFD数据格式,进一步通过XCMS软件包(参数设定fwhm=1,bw=2,snthersh=5,其余默认值)进行峰校正和峰积分[7],处理获得的矩阵导入Excel,对数据进行归一化处理,采用SIMCA-P 14.1软件进一步进行中心化和标准化处理,然后进行主成分分析和正交偏最小二乘法判别分析。通过S-plot载荷图寻找生物标志物,对差异代谢物做进一步双侧t检验分析(阿霉素组与正常组)和方差分析(P<0.05)。代谢物的鉴定通过标准质谱数据库(NIST数据库)检索匹配,并采用对照品对部分代谢物进行确认。
-
潜在生物标志物的代谢通路分析采用基于网页来源的Metaboanalyst(http://www.metaboanalyst.ca)软件进行分析[8]。
-
血清中LDH、CK-MB的测定结果和超声心动图如图1所示,与正常组相比,模型组血清LDH、CK-MB值明显升高,EF、FS值明显下降,表明造模成功,模型组小鼠心脏功能受损。经参附汤治疗后,与模型组相比,参附汤治疗组LDH、MB值明显降低(P<0.05),EF、FS值明显升高(P<0.05)。血清酶指标和超声心动图结果表明参附汤能够改善阿霉素引起的心功能损伤。
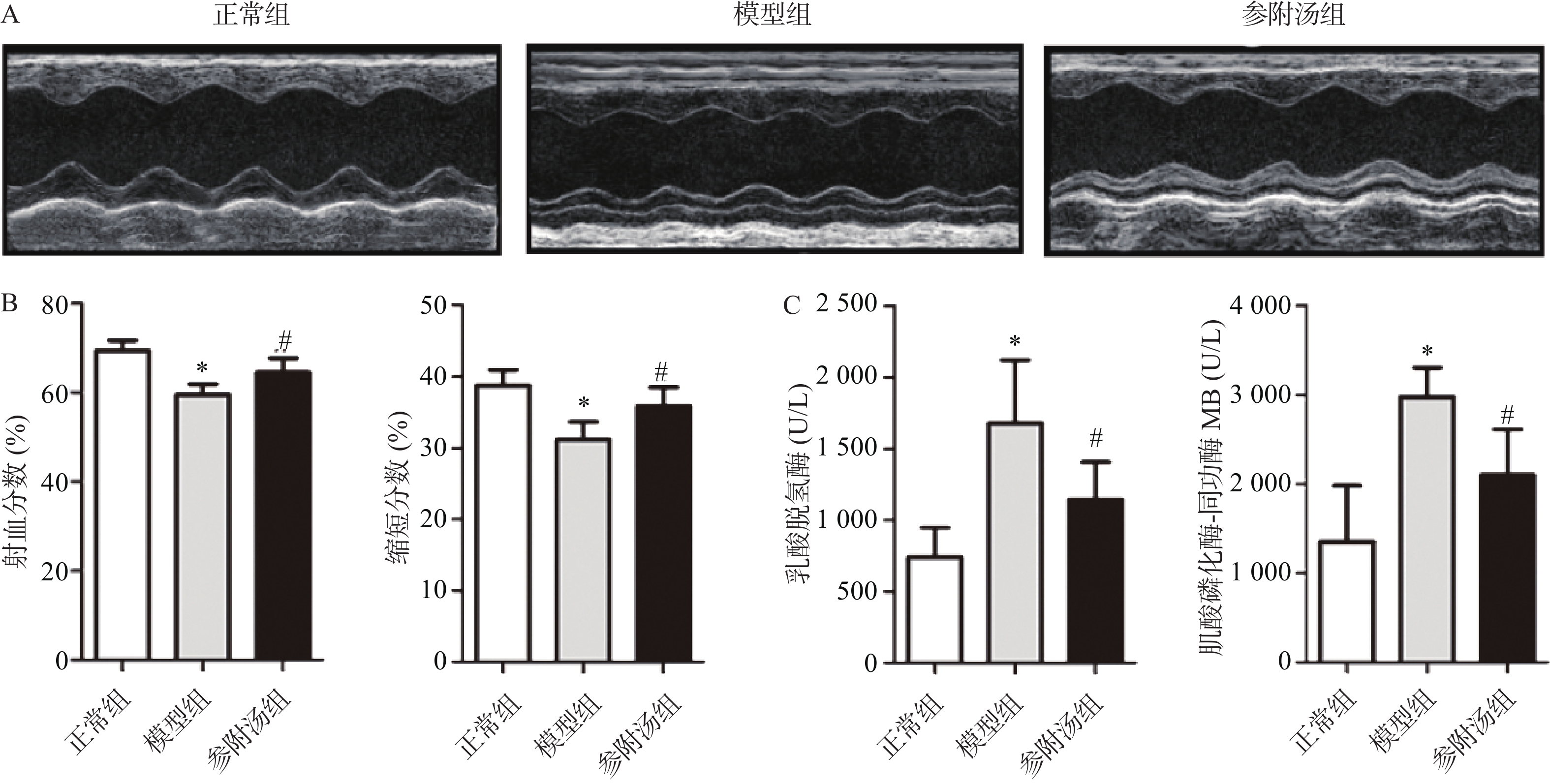
图 1 参附汤减轻阿霉素心肌损伤的超声心动图和生化分析结果(
$ \bar x \pm s $ ,n=8) -
采用QCs评价GC-MS系统的稳定性。整个分析过程一共进样9次QCs,不同组样品与QCs通过XCMS校正、积分、提取特征离子峰,一共得到352个离子峰,设定峰面积相对标准偏差小于30%提取变量,结果得到319个变量的数据集,占总变量数的90.6%。可见本研究建立的GC-MS分析系统稳定性良好。无监督的主成分分析结果如图2所示,可见9个QC紧密聚集在一起,这也进一步证实了分析系统具有良好的稳定性。
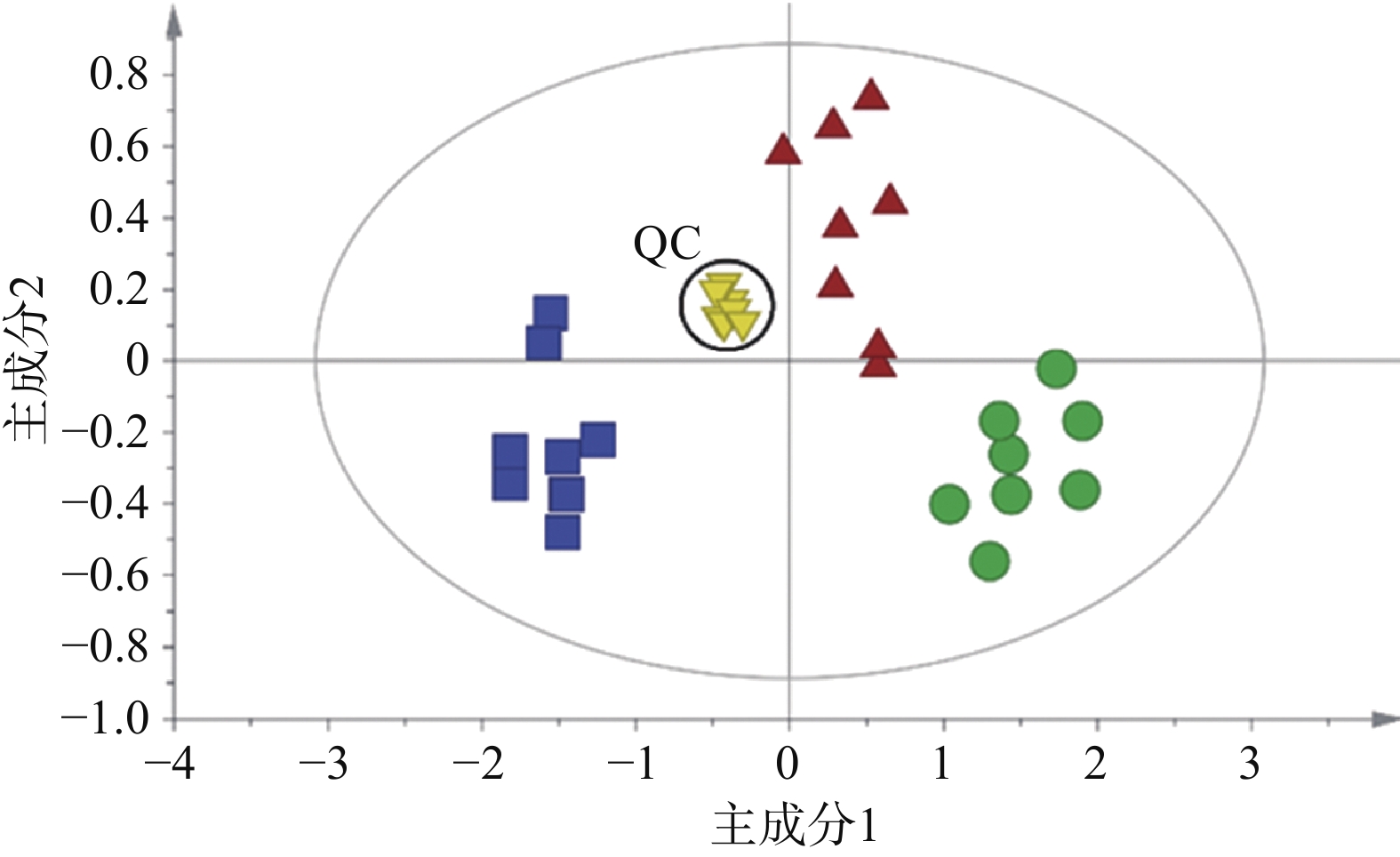
图 2 基于GC-MS数据集的主成分分析得分图
-
为研究小鼠心脏功能受损前后血清代谢物的差异与变化,并评估参附汤对小鼠受损心脏功能的保护作用,将实验获取的所有数据进行主成分分析。主成分分析能够将不同组别的样本区分开来,并且指示特征离子。将正常组和模型组的GC-MS数据进行正交偏最小二乘法判别分析,得出反映两组间离散程度的得分图(图3A)和寻找相关生物标志物的S-plot载荷图(图3B)。从得分图可以看出,正常组与模型组能实现明显分离,表明模型组小鼠心脏功能受损后,内源性物质代谢发生紊乱,血清代谢谱出现明显变化。S-plot载荷图能指引找到对2组分离贡献较大的代谢物,离原点越远贡献越大。利用S-plot载荷图,再结合峰面积相对强度的双侧t检验分析,一共鉴定了13种阿霉素心脏毒性潜在的生物标志物,如图4所示。

图 3 正交偏最小二乘法判别分析(A)和S-plot载荷图(B)

图 4 血清样品GC-MS分析总离子流图
在这13种潜在的生物标志物中,最常见的是氨基酸代谢失衡。模型组丙氨酸、苯丙氨酸水平升高,而异亮氨酸、缬氨酸水平降低。氨基酸是能量代谢的重要前体,能增加ATP的合成,另外苯丙氨酸是合成儿茶酚胺的重要原料,在应激状态下儿茶酚胺合成增多。其次是长链脂肪酸代谢受阻,模型组硬脂酸、油酸水平显著升高,而它们是正常心肌细胞能量代谢的主要原料。从三组的主成分分析得分图(图2)可以发现,参附汤治疗组与模型组、正常组之间均能分开,并且更加靠近正常组。此外,经参附汤治疗后,所鉴定的13种阿霉素心脏毒性潜在的生物标志物中的11种明显回调,如表1所示,这些结果与小鼠血清生化指标及超声心动图的改善相一致,表明参附汤能通过整体逆转小鼠血清代谢谱,从而发挥抗阿霉素诱导的心脏毒性的作用。
表 1 模型组与正常组的代谢物变化趋势及参附汤的回调作用(
$ \bar x \pm s $ ,n=8)序号 保留时间(min) 代谢物 基峰 m/z MS 碎片 峰相对强度 正常组 模型组 参附汤组 1 8.89 乳酸 73 191, 147, 117 (2.04±0.28)×10−1 (4.36±0.21)×10−1 ↑* (2.79±0.54)×10−1 ↓# 2 10.14 丙氨酸 116.1 190, 147, 73 (7.95±1.47)×10−3 (1.55±0.22)×10−2 ↑* (1.07±0.28)×10−2 ↓# 3 12.38 异亮氨酸 86.1 188, 147, 117, 73 (1.20±0.12)×10−3 (6.62±0.78)×10−4 ↓* (9.69±1.11)×10−4 ↑# 4 13.89 缬氨酸 144.1 218, 73 (2.02±0.41)×10−2 (1.03±0.11)×10−2 ↓* (1.71±0.39)×10−2 ↑# 5 17.04 琥珀酸 247.1 147, 75, 73 (6.45±0.90)×10−5 (1.11±0.23)×10−4 ↑* (7.02±1.62)×10−5 ↓# 6 22.87 苹果酸 73 233, 189, 147 (8.88±1.19)×10−4 (1.67±0.20)×10−3 ↑* (1.04±0.26)×10−3 ↓# 7 24.03 苯丙氨酸 120.1 146, 91, 73 (8.79±1.07)×10−4 (1.57±0.20)×10−3 ↑* (1.15±0.15)×10−3 ↓# 8 30.84 甘油-3-磷酸 299.1 357, 147, 73 (7.95±0.83)×10−5 (1.25±0.18)×10−4 ↑* (1.08±0.14)×10−4 ↓ 9 32.25 柠檬酸 273.11 147, 73 (5.00±0.64)×10−4 (3.04±0.53)×10−4 ↓* (4.33±0.56)×10−4 ↑# 10 33.36 酪氨酸 179.1 208, 73 (1.70±0.30)×10−3 (1.04±0.17)×10−3 ↓* (1.25±0.31)×10−3 ↑ 11 40.85 油酸 73.01 339, 129, 117 (2.22±0.55)×10−3 (3.61±0.63)×10−3 ↑* (2.51±0.51)×10−3 ↓# 12 41.34 硬脂酸 117 341, 145, 129 (1.76±0.63)×10−3 (3.00±0.40)×10−3 ↑* (2.11±0.45)×10−3 ↓# 13 43.10 花生四烯酸 93.1 133, 117, 91 (1.57±0.11)×10−4 (2.48±0.33)×10−4 ↑* (1.72±0.28)×10−4 ↓# 注:*P<0.05,与正常组比较;#P<0.05,与模型组比较 -
采用Metaboanalyst软件对逆转代谢标志物在参附汤治疗组与阿霉素组的相对峰面积进行代谢通路分析,以代谢通路重要性>0.1和统计学分析P<0.05为靶向通路筛选界值,结果如图5所示。表明苯丙氨酸、酪氨酸和色氨酸的合成、花生四烯酸代谢、苯丙氨酸代谢、三羧酸循环和二羧酸代谢通路是参附汤主要的靶向代谢通路。其中苯丙氨酸、酪氨酸和色氨酸是蛋白质代谢的重要成分,花生四烯酸代谢是炎症调控代谢途经,三羧酸循环、二羧酸代谢是心肌能量代谢的重要途经。表明参附汤可以通过改善心肌蛋白质代谢、能量代谢和减轻炎症效应,从而发挥抗阿霉素心脏毒性的作用。
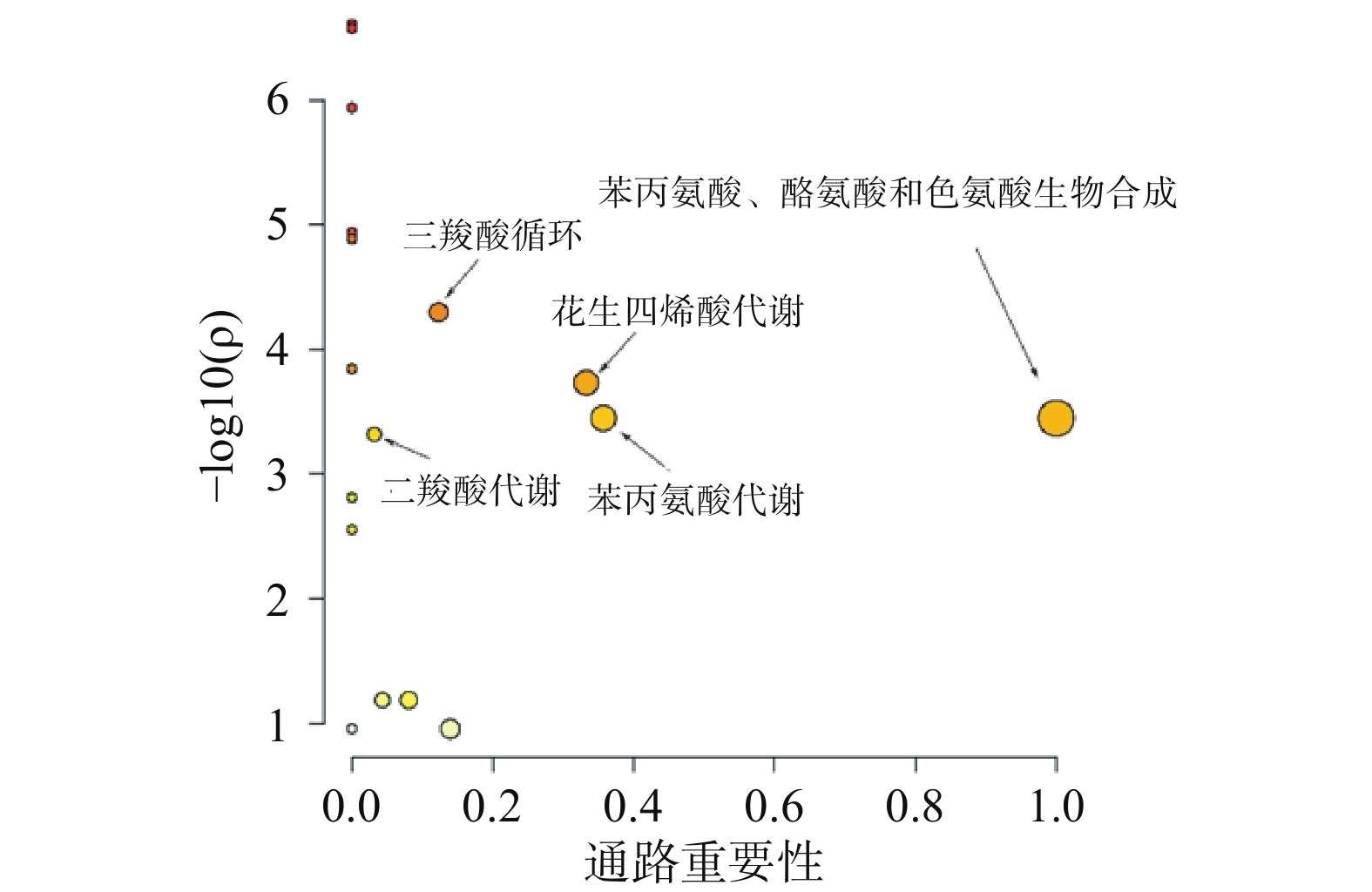
图 5 参附汤治疗组与模型组差异代谢物的通路分析
-
本研究建立了一种基于GC-MS的代谢组学研究方法,用于探索阿霉素对小鼠心脏毒性以及参附汤抗阿霉素心脏毒性的作用机制。单变量和多变量统计学分析结果提示正常组、模型组和参附汤治疗组的代谢谱明显不同,一共鉴定出13种潜在阿霉素心脏毒性血清生物标志物,以这些生物标志物为潜在的药物靶标,参附汤能够明显逆转其中11种代谢物,表明参附汤对阿霉素所致的心功能损伤有治疗作用。多变量统计学分析代谢通路表明,苯丙氨酸、酪氨酸和色氨酸的合成、花生四烯酸代谢、苯丙氨酸代谢、三羧酸循环和二羧酸代谢是参附汤主要的靶向代谢通路。
Serum metabolomic study of Shenfu decoction on adriamycin-induced cardiomyopathy in mice
-
摘要:
目的 应用血清代谢组学技术,研究参附汤对阿霉素心脏毒性的逆转作用,探讨其作用机制。 方法 建立阿霉素致心肌病BALB/c小鼠模型,给予相应的干预,测定血清乳酸脱氢酶(LDH)、肌酸磷化酶-同功酶MB(CK-MB);超声心动图测定射血分数(EF)和缩短分数(FS)。采集小鼠血清进行气相色谱-质谱(GC-MS)分析,所得数据经多变量和单变量统计分析,比较正常组、模型组和参附汤治疗组小鼠血清中内源性代谢物的变化,寻找参附汤逆转阿霉素心脏毒性潜在的生物标志物,采用代谢通路分析探讨参附汤靶向代谢通路。 结果 模型组血清LDH、CK-MB水平明显升高,EF、FS值明显下降,表明造模成功,经参附汤治疗后上述指标显著改善。代谢组学分析鉴定出13种阿霉素对心脏毒性的潜在生物标志物,参附汤对其中11种代谢物具有显著的逆转作用。代谢通路分析表明,苯丙氨酸、酪氨酸和色氨酸的合成、花生四烯酸代谢、苯丙氨酸代谢、三羧酸循环和二羧酸代谢是参附汤主要的靶向代谢通路。 结论 参附汤能通过调节失衡的苯丙氨酸、酪氨酸和色氨酸的合成,以及花生四烯酸、苯丙氨酸、二羧酸代谢和三羧酸循环发挥逆转阿霉素心脏毒性的作用。 Abstract:Objective To study the reversal effect of Shenfu decoction(SFD)on adriamycin-induced cardiomyopathy and explore its mechanism by using serum metabolomic technology. Methods The BALB/c mouse model of cardiomyopathy induced by adriamycin was established. The corresponding intervention was given. The serum lactate dehydrogenase(LDH)and creatine phosphatase isoenzyme MB(CK-MB)were measured. The ejection fraction (EF) and shortening fraction (FS) were measured by echocardiography. Mouse serum was collected for gas chromatography-mass spectrometry (GC-MS) analysis. The data obtained was analyzed by multivariate and univariate statistical analysis to compare the changes of endogenous metabolites in the serum of mice in the normal group, model group and Shenfu decoction treatment group, to find the potential biomarkers of Shenfu decoction to reverse the adriamycin-induced cardiomyopathy. Metabolic pathway analysis was used to explore the targeted metabolic pathway of Shenfu decoction. Results The levels of serum LDH and CK-MB in the model group were increased significantly, and the values of EF and FS decreased significantly, indicating that the model was successfully established. The above indicators were significantly improved after treatment with Shenfu decoction. 13 potential biomarkers of adriamycin-induced cardiomyopathy were identified by metabonomic analysis, and Shenfu decoction had significant reversal effect on 11 metabolites. Metabolic pathway analysis showed that the synthesis of phenylalanine, tyrosine and tryptophan, arachidonic acid metabolism, phenylalanine metabolism, tricarboxylic acid cycle and dicarboxylic acid metabolism were the main targeted metabolic pathways of Shenfu decoction. Conclusion Shenfu decoction can reverse adriamycin-induced cardiomyopathy by regulating the unbalanced synthesis of phenylalanine, tyrosine and tryptophan, as well as the metabolism of arachidonic acid, phenylalanine, dicarboxylic acid and tricarboxylic acid cycle. -
Key words:
- adriamycin /
- cardiomyopathy /
- Shenfu decoction /
- GC-MS /
- metabolomics
-
神经损伤是世界范围内致残和导致死亡的主要原因,神经损伤疾病患病率的大幅增加导致了全社会的健康负担加重[1]。神经损伤主要包括颅脑损伤(TBI)和缺血性脑卒中(CI)等,其中TBI是最常见的神经损伤类型[2],分为急性和慢性两个阶段,炎症是这两个阶段的共同特征,目前还没有有效治疗TBI的药物和方法,迫切需要寻找具有更广泛作用的药物,以缓解TBI后炎症反应[3]。脑缺血是神经损伤患者死亡的最常见病因之一[4],大脑因供血中断而致脑缺血,进而导致中风等神经损伤性疾病[5]。CI诱导了多种细胞死亡形式,如兴奋性毒性、酸毒性和离子失衡、氧化/氧化应激、炎症[6]、凋亡和梗死周围去极化等。目前CI最有效的治疗手段是静脉溶栓和血管内取栓以达到快速再灌注,这两者都能降低患者致残率,但都需要在发病4小时内尽快完成[4],目前尚无有效的药物治疗CI患者。
中药具有多途径、多靶点的特点,已有2000多年的发展历史和临床用药经验[7],如唐·孙思邈所著《备急千金方》中“小续命汤治卒中风欲死”和“大秦艽汤(金·刘完素)治中风”等。因此,阐明在临床上广泛使用的中药的作用机制是本文关注的重点。益母草来自唇形科植物益母草(Leonurus japonicus Houtt)的新鲜或干燥地上部分,临床上主要用于子宫收缩和镇静[8]。现代药理学研究发现其具有子宫收缩、抗炎、镇痛和抗氧化作用等[9]。值得引起关注的是,益母草对神经损伤也有保护作用[10]。然而,关于益母草治疗神经损伤的物质基础和作用机制的研究还未见报道。因此有必要阐明益母草治疗神经损伤的物质基础和作用机制。
本研究旨在利用网络药理学预测益母草的活性成分、靶点及相关通路来探讨其治疗神经损伤的潜在分子机制,为益母草的药理机制深入研究和临床应用提供参考。
1. 材料与方法
1.1 数据库及软件
中药系统药理学分析平台(TCMSP)和中药分子机制的生物信息学分析工具(BATMAN-TCM);活性成分靶标预测数据库(SwissTargetPrediction, STP);人类基因注释数据库(GeneCards);疾病靶点标准化数据库(Uniprot);京都基因与基因组百科全书; 蛋白-蛋白相互作用网络平台(STRING 11.0);Venny2.1软件、Cytoscape 3.6.0软件和在线作图工具微生信。
1.2 益母草活性成分的筛选
通过TCMSP和BATMAN-TCM数据库输入“yimucao”,搜索得到益母草的活性成分, 然后,在TCMSP中设置口服生物利用度(OB)≥30%及药物相似性(DL)≥0.18;在BATMAN-TCM中设置“药物-靶点”相似性模型阀值≥20,调节P值≤0.05,筛选活性成分。
1.3 活性成分靶点的预测和筛选
在获得益母草活性成分的基础上,检索TCMSP和STP数据库,限定种属为“Homo sapiens(人类)”,获取活性成分的作用靶点。通过PubChem数据库来确证收集到的活性成分,将其标准化并下载 SMILES序列。再通过TCMSP数据库搜索确证后的活性成分的靶点,将搜索的靶点按照度值从大到小排列后得到益母草的潜在靶点。在数据库STP中,搜索SMILE式,筛选条件为“概率>0”,删除重复值后预测得到药物的潜在靶点。此外,由于益母草中葫芦巴碱已被证实具有较好的治疗神经损伤作用,故将该化合物也纳入活性成分范围内[11],并通过TCMSP和STP数据库获取其成分靶点。
1.4 疾病相关靶点的筛选
通过GeneCards、DisGenet、OMIM数据库以疾病名称“cerebral ischemia”和“traumatic brain injury”进行检索,获得神经损伤相关靶标。
1.5 药物靶点与疾病靶点标准化及PPI网络构建
利用疾病靶点标准化数据库Uniprot,分别上传上述得到的益母草潜在靶点与神经损伤相关靶点名,获取其靶点的标准基因名以及Uniprot ID。为明确益母草治疗神经损伤疾病的潜在靶点,将二者的靶点上传至Venny 2.1 软件绘制韦恩图,并导出交集的基因。再将筛选得到的共有靶标蛋白上传至STRING平台,选择“multiple proteins”模式,建立药物靶蛋白-疾病靶蛋白相互作用网络,结合分值取中等“medium confidence(≥0.4)”,其余参数默认,利用 Cytoscape3.6.0软件构建益母草治疗神经损伤的PPI网络。利用cytoHubba插件计算 PPI网络每个节点的度值,筛选益母草治疗神经损伤的核心靶点。
1.6 富集分析
在上述STRING中的结果下,选择“Analysis”,点击下载“Biological Process(GO)”“Molecular Function(GO)”“Cellular Component(GO)”“KEGG Pathways”。阈值设置为P≤0.01, 并按照涉及的靶点数目多少进行排序,得到GO气泡图和 KEGG 信号通路条形图。
2. 结果
2.1 益母草活性成分的筛选
通过TCMSP数据库检索到益母草已报道的化学成分,以ADME参数OB≥30%、DL≥0.18进行筛选,得到益母草活性成分8个;BATMAN-TCM数据库检索到10个益母草活性成分;加上文献检索的1个化合物共19个(表1)。再将这19个活性成分输入TCMSP以及STP数据库,搜索的结果经过筛选去除重复项后共得到654个益母草潜在靶点。
表 1 益母草筛选所得活性成分序号 化合物 来源数据库 1 没食子酸 BATMAN-TCM 2 水苏糖 BATMAN-TCM 3 芦丁 BATMAN-TCM 4 月桂酸 BATMAN-TCM 5 水苏碱 BATMAN-TCM 6 益母草素 BATMAN-TCM 7 西班牙夏罗草酮 BATMAN-TCM 8 鸟嘌呤 BATMAN-TCM 9 益母草碱 BATMAN-TCM 10 4-胍基丁醇 BATMAN-TCM 11 鼬瓣花二萜 TCMSP 12 ZINC04073977 TCMSP 13 前益母草二萜 TCMSP 14 异前益母草二萜 TCMSP 15 槲皮苷 TCMSP 16 花生四烯酸 TCMSP 17 异鼠李素 TCMSP 18 山奈酚 TCMSP 19 葫芦巴碱 文献 2.2 疾病靶点的筛选
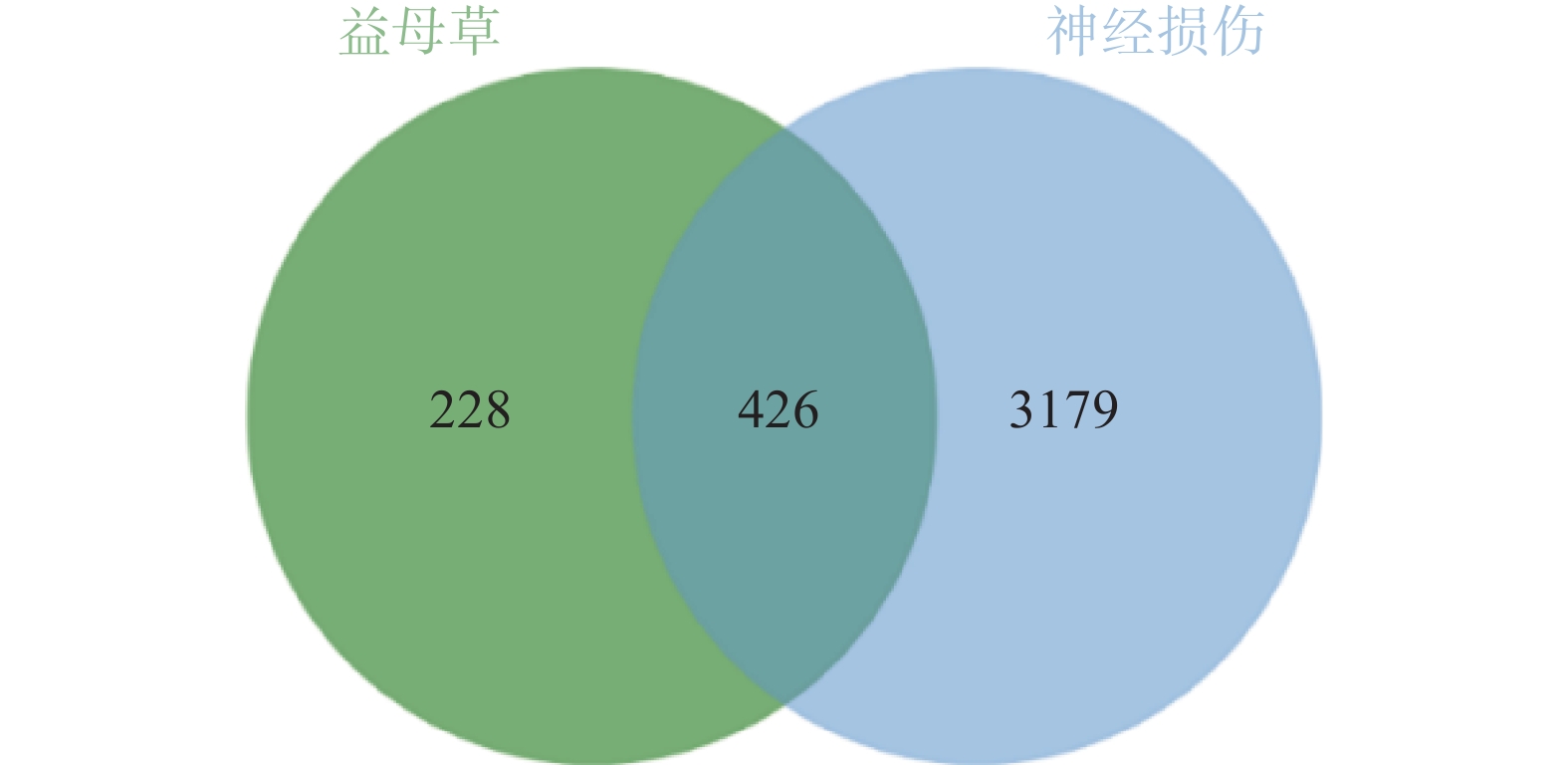
通过GeneCards、DisGenet和OMIM数据库以疾病名称为“cerebral ischemia”和“traumatic brain injury”分别进行检索,在DisGenet数据库中没有检测到TBI靶点,删除重复值后,得到神经损伤的靶点3605个,将疾病相关的靶点与益母草靶点进行Venn交集分析,筛选得到益母草治疗神经损伤的潜在靶点426个,并获得药物-疾病共同靶点基因韦恩图(图1)。
2.3 PPI 网络分析
将益母草治疗神经损伤的426个潜在靶点,导入STRING数据库,将相互作用靶点的结果导入Cytoscape 3.6.0进行可视化分析,得到由331个节点、6955条边共同组成的网络(图2),同时得到网络中关键靶点的度值(表2)。如图2所示,与神经损伤相关度较高的靶点(度值≥139)为丝氨酸/苏氨酸蛋白激酶1(AKT1)、白细胞介素6(IL-6)受体、血管内皮生长因子A(VEGFA)、半胱氨酸蛋白酶3(CASP3)、肿瘤蛋白P53(TP53)、基质金属蛋白酶-9 (MMP9)。度值大的靶点提示在网络调控中起关键作用,这些度值大的靶点很可能是益母草治疗神经损伤的关键靶点。
表 2 益母草治疗神经损伤相关靶点信息基因 度值 靶点名称 数据库中代码 AKT1 225 丝氨酸/苏氨酸蛋白激酶 P31749 IL6 217 白介素6 P05231 VEGFA 196 血管内皮生长因子A P15692 TNF 187 肿瘤生长因子 P01375 TP53 186 细胞肿瘤抗原P53 P04637 SRC 165 原癌基因酪氨酸受体激酶 P12931 CASP3 163 胱天蛋白酶-3 P42574 MAPK1 160 丝裂原活化蛋白激酶1 P28482 CXCL8 157 白介素8 P10145 EGFR 153 表皮生长因子受体 P00533 EGF 150 前表皮生长因子 P01133 PTGS2 146 牛前列腺素G/H合成酶2 P35354 MAPK8 146 丝裂原活化蛋白激酶8 P45983 MYC 146 原癌基因蛋白Myc P01106 JUN 145 转录因子AP-1 P05412 STAT3 143 信号传导及转录激活子3 P40763 FOS 143 原癌基因c-Fos P01100 MMP9 139 基质金属蛋白酶9 P14780 IL-1β 133 白介素1β P01584 2.4 GO生物过程和KEGG通路富集分析
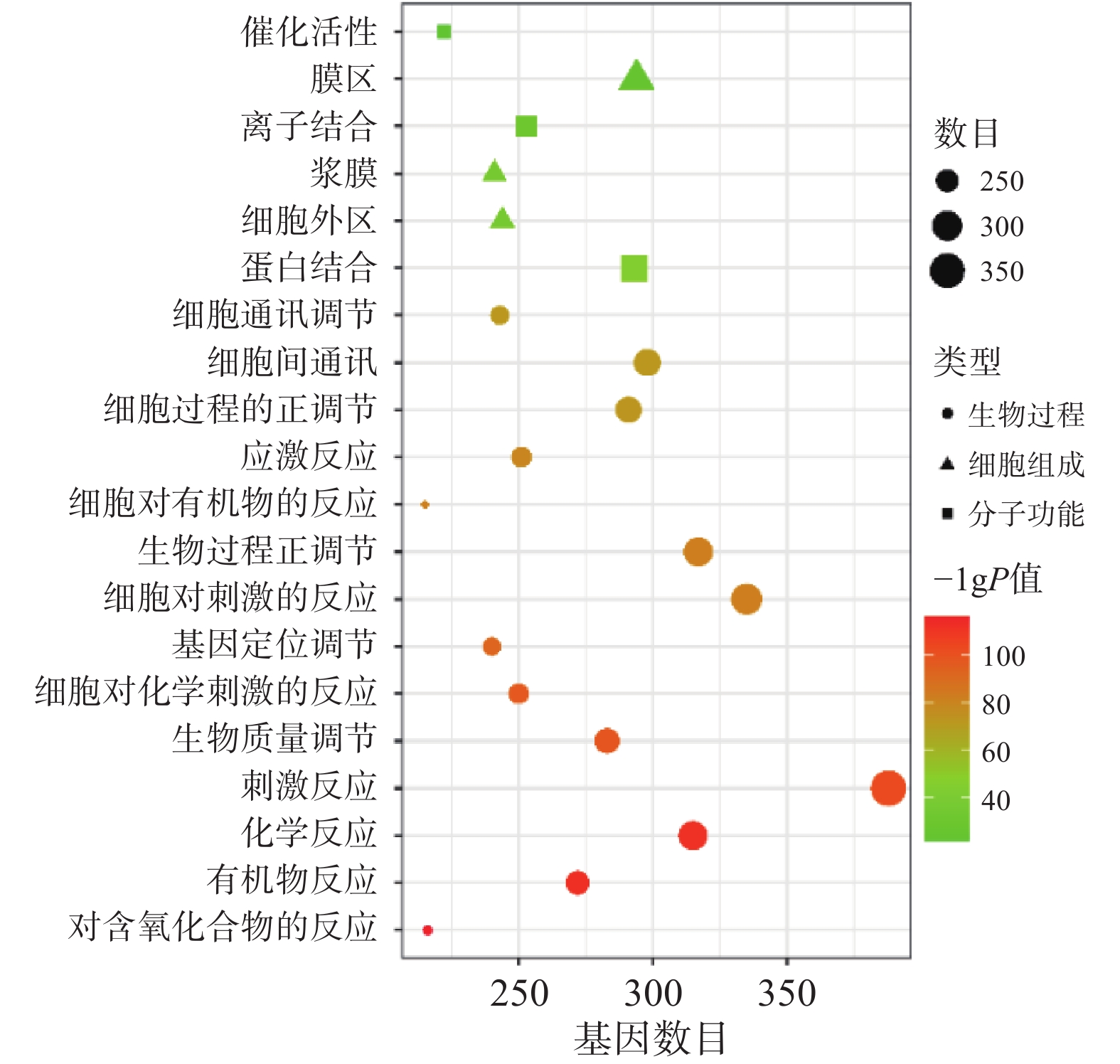
将“2.2” 项下获取的426个潜在的治疗神经损伤的靶点通过STRING进行生物过程(BP)、细胞组分(CC)和分子功能(MF)分析,以 P<0.01为条件,筛选靠前的GO富集分析,如图3所示。图中纵坐标表示富集条目,横坐标表示基因计数,颜色深浅代表-log10(p)值大小。其中 GO-BP 主要为应激反应、生物调节和细胞通讯等;GO-CC主要为细胞膜等;GO-MF主要为蛋白质结合、离子结合和催化还原活性等。
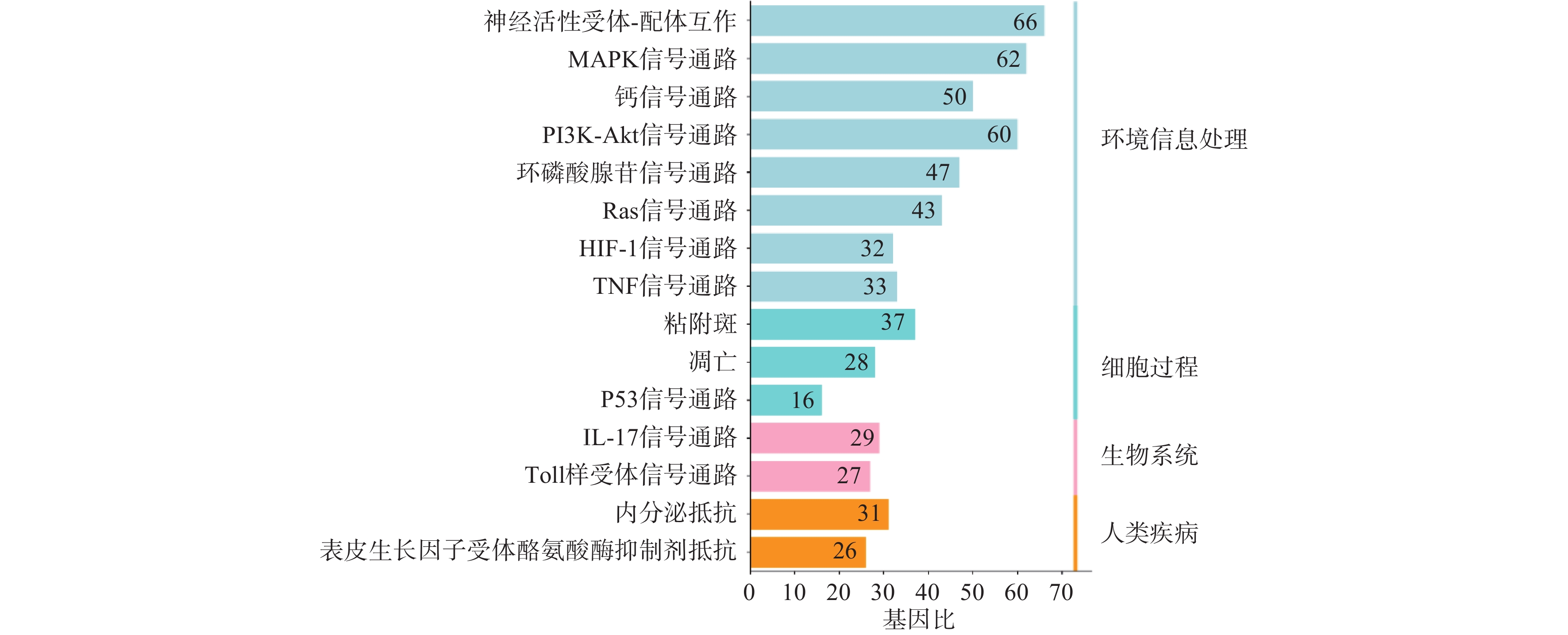
利用STRING数据库对益母草治疗神经损伤的潜在的426个靶点进行富集分析,筛选出显著的前16条信号通路(P<0.01),主要涉及的信号通路为MAPK、Toll样受体、PI3K-Akt、肿瘤坏死因子、IL-17和凋亡等信号通路(图4)。
2.5 药物“活性成分-靶点”网络的构建与分析
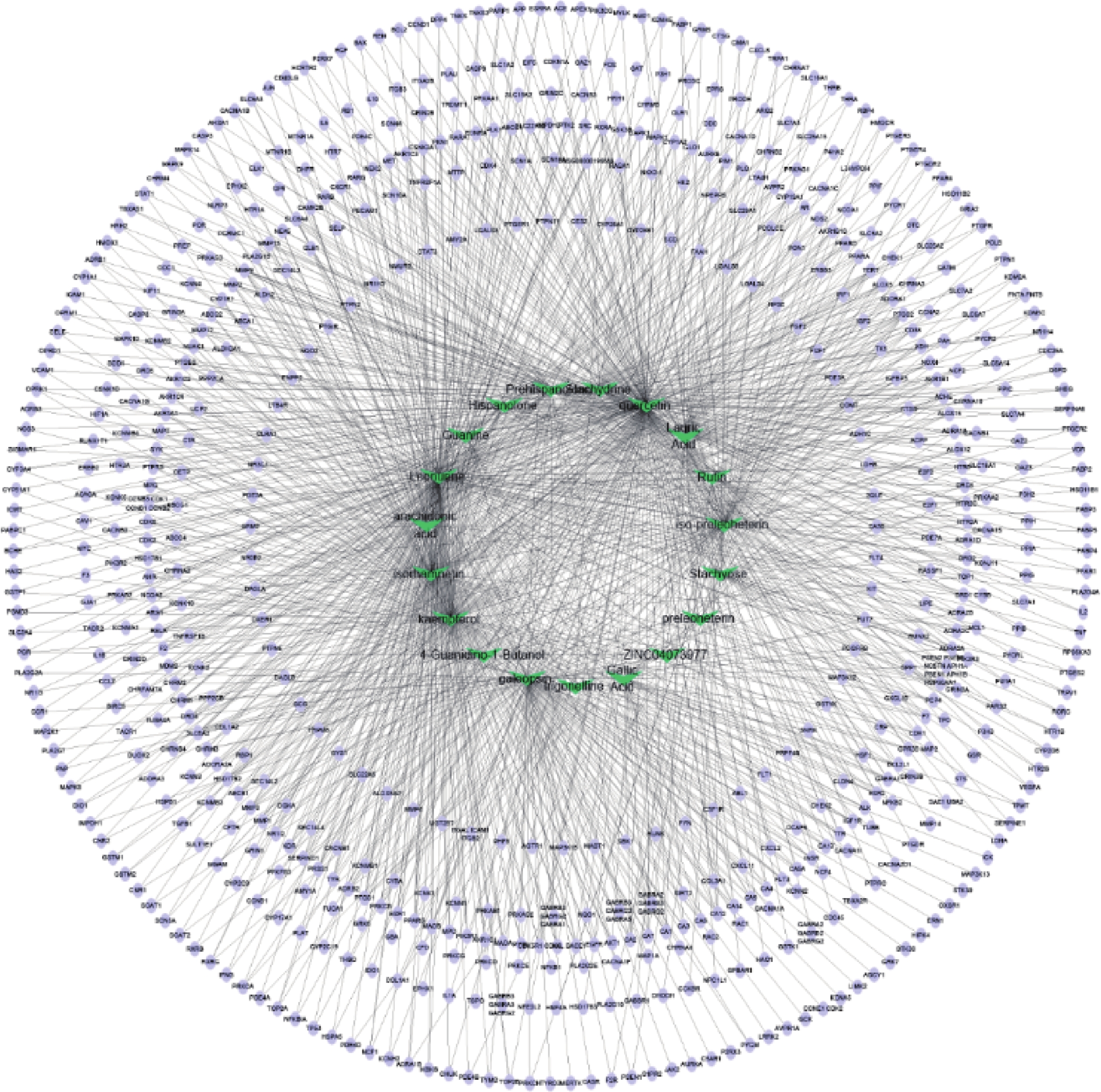
通过Cytoscape3.6.0软件得到药物“活性成分-靶点”的网络(图5)。网络中化合物19个,靶点426个。在图5中,箭头节点代表益母草中化合物,椭圆形节点代表靶点,结果可以明显看出益母草中度值相对较高的化合物有槲皮素、益母草碱、山奈酚、异鼠李素、水苏碱、葫芦巴碱等,这些化合物可能是益母草治疗神经损伤的关键化合物。
3. 讨论
本研究采用网络药理学方法,借助相关数据库以及各种绘图软件对益母草治疗神经损伤的物质基础和作用机制进行研究。共筛选出益母草有效活性成分19个,对应活性成分靶点654个。其中,益母草与神经损伤的共同基因有426个,“药物活性成分-共同靶点”网络与PPI网络结合分析,结果发现,益母草治疗神经损伤的关键活性成分有槲皮素、益母草碱、山奈酚、异鼠李素、水苏碱、葫芦巴碱等,其中槲皮素对脑缺血的作用最为突出。槲皮素通过发挥抗氧化[12]、抗炎和抗凋亡作用[13]对脑缺血的病理学改变产生了积极的治疗作用。益母草碱通过抗氧化、抗凋亡、保护线粒体和激活Nrf-2/HO-1信号通路发挥血脑屏障保护作用[14]。山萘酚具有抗氧化、抗炎、抗癌和预防心血管疾病等多种药理活性[15]。异鼠李素具有保护心脑血管、抗肿瘤、抗炎、抗氧化、保护器官、预防肥胖等作用[16]。水苏碱通过多种分子机制(包括TGF-β、ers介导的细胞凋亡、MMPs/TIMPs、NF-κB和JAK/STAT)抑制细胞外基质(ECM)沉积、降低炎症和氧化应激,以及血管生成保护血管等发挥神经保护作用[17]。葫芦巴碱具有抗凋亡、抗炎、抗氧化、神经保护等多种药理作用,具有改善认知能力的潜力。
根据益母草靶蛋白互作网络图分析可知,益母草治疗神经损伤的核心靶点主要为:AKT1,IL-6,VEGFA,CASP3, TP53,MMP-9。AKT1能够通过丝氨酸和/或苏氨酸磷酸化介导调节细胞代谢、增殖、细胞存活、胰岛素信号传导、生长和血管生成过程。据报道[18]阿托伐他汀通过抑制JNK3/cJun/caspase-3,增强Akt-nNOS信号通路,抑制脑缺血大鼠脑内细胞凋亡,对脑缺血再灌注有保护作用。IL-6可调节多种细胞的生长与分化,具有调节免疫应答、急性期反应及造血功能,并在机体的抗感染免疫反应中起重要作用。IL-6作为促炎细胞因子在脑缺血中的作用可能是通过NF-κB通路来实现的[19]。据报道,LncRNA MEG8通过miR-130a-5p/VEGFA信号靶向减轻缺血性中风后的脑缺血[20],通过靶向VEGFA,下调microRNA-195促进血管生成[21]。Caspase-3(CASP3)是细胞凋亡途径中最关键的酶类之一,与癌症的发生、衰老、心脑血管疾病的发生等有着重要联系。Nahid等[22]研究发现通过降低Bax/Bcl-2比值和caspase-3活化,可减轻脑缺血后海马CA1神经元损伤,改善脑缺血损伤引起的功能和记忆丧失。TP53是神经元凋亡的主要调节因子,任何降低TP53稳定性及其向线粒体迁移的方法都可以减轻缺血性脑区的神经元损失[23]。MMP-9是一种明胶酶,大脑中许多细胞都能分泌MMP-9。MMP-9降解细胞外基质成分,从而引发中风,Zinnhardt等[24]研究发现脑缺血的发生会促进基质金属蛋白酶(MMPs)的产生,尤其是MMP-9,另外MMP-9的激活又可引起血脑屏障受损。
GO功能富集分析发现,益母草治疗神经损伤的基因功能主要体现在生物调节、氧化应激反应、细胞通讯等生物学过程以及蛋白质结合、离子结合和催化还原等。KEGG信号通路富集分析显示,益母草治疗神经损伤所涉及的TNF信号通路、MAPK信号通路、TP53信号通路、PI3K-Akt信号通路的P值较小,被显著富集。TNF具有促进细胞生长、分化、凋亡及诱发炎症等生物学效应。TNF-α可以激活JNK,Caspase蛋白酶和转录因子NF-kB这三条信号通路,实现其免疫调节和细胞凋亡的生物学功能,从而对脑缺血产生影响。级联p38-MAPK的转导通路位于中枢神经系统,在缺血、缺氧等条件刺激下可被激活。可通过p38 MAPK和c-Jun抑制炎症反应,对损伤后的神经有保护作用[25]。Yao等[26]发现通过抑制MAPK信号通路的激活,恢复神经功能,减轻血脑屏障通透性破坏,对脑缺血产生保护作用。缺血神经元释放的内源性配体激活TLR信号通路,导致大量炎症细胞因子TNF-α、IL-1β、iNOS的产生,从而引起脑缺血后继发性炎症损伤。TLRs介导的缺血耐受可作为预防和治疗脑缺血的重要靶点[27]。TP53是一个肿瘤抑制蛋白,调节各种各样基因的表达,包括细胞凋亡等,此外TP53可不依赖其活性,仅作为一个转录因子来引发凋亡通路。抑制NF-κB及下游TP53可显著减轻神经元自噬和凋亡,具有显著的神经保护作用。NF-κB、TP53及其介导的自噬和凋亡在脑缺血再灌注损伤恶化中也起关键作用[28]。PI3K/AKT信号通路是一条与增殖,分化和凋亡相关的信号通路[29]。
综上所述,本研究应用网络药理学的方法预测了益母草治疗神经损伤的主要活性成分和潜在分子机制,但由于所使用平台的数据收录,更新相对滞后,中药活性成分筛选条件口服生物利用度与类药性并不是唯一的标准,因而,研究预测的结果有其局限性。需要在今后的实验研究中进一步阐明和验证益母草中活性成分的作用靶点,从而完善其治疗神经损伤的有效化学成分及作用机制。
-
图 1 参附汤减轻阿霉素心肌损伤的超声心动图和生化分析结果(
$ \bar x \pm s $ ,n=8)注:A. 正常组、模型组、参附汤组典型的M-型超声心动图;B. 射血分数和缩短分数分析结果;C. 血清乳酸脱氢酶和肌酸磷化酶-同功酶MB测定结果*P<0.05,与正常组比较;#P<0.05,与模型组比较
表 1 模型组与正常组的代谢物变化趋势及参附汤的回调作用(
$ \bar x \pm s $ ,n=8)序号 保留时间(min) 代谢物 基峰 m/z MS 碎片 峰相对强度 正常组 模型组 参附汤组 1 8.89 乳酸 73 191, 147, 117 (2.04±0.28)×10−1 (4.36±0.21)×10−1 ↑* (2.79±0.54)×10−1 ↓# 2 10.14 丙氨酸 116.1 190, 147, 73 (7.95±1.47)×10−3 (1.55±0.22)×10−2 ↑* (1.07±0.28)×10−2 ↓# 3 12.38 异亮氨酸 86.1 188, 147, 117, 73 (1.20±0.12)×10−3 (6.62±0.78)×10−4 ↓* (9.69±1.11)×10−4 ↑# 4 13.89 缬氨酸 144.1 218, 73 (2.02±0.41)×10−2 (1.03±0.11)×10−2 ↓* (1.71±0.39)×10−2 ↑# 5 17.04 琥珀酸 247.1 147, 75, 73 (6.45±0.90)×10−5 (1.11±0.23)×10−4 ↑* (7.02±1.62)×10−5 ↓# 6 22.87 苹果酸 73 233, 189, 147 (8.88±1.19)×10−4 (1.67±0.20)×10−3 ↑* (1.04±0.26)×10−3 ↓# 7 24.03 苯丙氨酸 120.1 146, 91, 73 (8.79±1.07)×10−4 (1.57±0.20)×10−3 ↑* (1.15±0.15)×10−3 ↓# 8 30.84 甘油-3-磷酸 299.1 357, 147, 73 (7.95±0.83)×10−5 (1.25±0.18)×10−4 ↑* (1.08±0.14)×10−4 ↓ 9 32.25 柠檬酸 273.11 147, 73 (5.00±0.64)×10−4 (3.04±0.53)×10−4 ↓* (4.33±0.56)×10−4 ↑# 10 33.36 酪氨酸 179.1 208, 73 (1.70±0.30)×10−3 (1.04±0.17)×10−3 ↓* (1.25±0.31)×10−3 ↑ 11 40.85 油酸 73.01 339, 129, 117 (2.22±0.55)×10−3 (3.61±0.63)×10−3 ↑* (2.51±0.51)×10−3 ↓# 12 41.34 硬脂酸 117 341, 145, 129 (1.76±0.63)×10−3 (3.00±0.40)×10−3 ↑* (2.11±0.45)×10−3 ↓# 13 43.10 花生四烯酸 93.1 133, 117, 91 (1.57±0.11)×10−4 (2.48±0.33)×10−4 ↑* (1.72±0.28)×10−4 ↓# 注:*P<0.05,与正常组比较;#P<0.05,与模型组比较  下载: 导出CSV
下载: 导出CSV
-
[1] BHAGAT A, KLEINERMAN E S. Anthracycline-induced cardiotoxicity: causes, mechanisms, and prevention[J]. Adv Exp Med Biol,2020,1257:181-192. [2] 范颖, 才丽平, 于彩娜, 等. 参附汤、芪附汤、姜附汤对阿霉素心脏毒性损伤大鼠线粒体途径细胞凋亡的影响[J]. 中国实验方剂学杂志, 2010, 16(8):135-138. doi: 10.3969/j.issn.1005-9903.2010.08.042 [3] 王雪梅, 刘佳, 付殿斌, 等. 参附汤萃取液成分对阿霉素致心衰大鼠血流动力学、心肌自噬及凋亡的影响[J]. 陕西中医学院学报, 2014, 37(4):75-78. [4] YAN X, WU H J, REN J X, et al. Shenfu Formula reduces cardiomyocyte apoptosis in heart failure rats by regulating micro-RNAs[J]. J Ethnopharmacol,2018,227:105-112. doi: 10.1016/j.jep.2018.05.006 [5] WANG M, CHEN L, LIU D, et al. Metabolomics highlights pharmacological bioactivity and biochemical mechanism of traditional Chinese medicine[J]. Chem Biol Interact,2017,273:133-141. doi: 10.1016/j.cbi.2017.06.011 [6] WU G S, LI H K, ZHANG W D. Metabolomics and its application in the treatment of coronary heart disease with traditional Chinese medicine[J]. Chin J Nat Med,2019,17(5):321-330. [7] DOMINGO-ALMENARA X, SIUZDAK G. Metabolomics data processing using XCMS[J]. Methods Mol Biol,2020,2104:11-24. [8] CHONG J, XIA J G. Using MetaboAnalyst 4.0 for metabolomics data analysis, interpretation, and integration with other omics data[J]. Methods Mol Biol,2020,2104:337-360. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3139
- HTML全文浏览量: 1304
- PDF下载量: 24
- 被引次数: 0
