

 下载:
下载:
 下载:
下载:

-
海洋生物毒素是一类由海洋生物产生的天然活性物质,具有化学结构新颖,生物活性高,作用于钠、钾、钙等离子通道等特点,有望开发为先导化合物,为新型海洋药物的研发提供新策略,但其强烈毒性和高致死率也对人类健康产生巨大威胁[1]。目前,药物毒性评价多采用细胞模型和动物模型,由于细胞模型不能重现体内真实环境,动物模型存在伦理和物种差异等问题,近年来,在微流控技术基础上发展起来的器官芯片模型,可以模拟体内组织器官的结构和功能,再现体内微环境,在药物毒性评价与检测领域展现出极大潜力。本文构建了一种可模拟心血管结构与功能的心血管芯片模型,用于四种典型的深海毒素,即大田软海绵酸(OA)、芋螺毒素(CTX)、河豚毒素(TTX)和环亚胺毒素(GYM)的致伤评价,并初步探索了雷公藤甲素对毒素致伤的保护作用,旨在为深海毒素的防护提供参考。
-
SC-1B匀胶机、BP-2B型烘胶台(北京创世威纳科技有限公司);JKG-2A光刻机(上海学泽光学机械有限公司);PDC-32G-2型等离子清洗机(美国Harrick公司);CKX41型高倍显微镜(日本OLYMPUS公司);Harris Uni-Corel 15072型打孔器(美国Harris公司);超净工作台、CO2细胞培养箱、离心机、超低温冰箱(美国Thermo Fisher公司);Synergy 4多功能酶标仪(美国Bio-TEK公司);Harvard蠕动泵p-70(美国Harvard仪器公司);十万分之一电子分析天平(日本A&D公司)。
聚二甲基硅氧烷(PDMS)预聚剂和固化剂(美国Dow Corning公司);硅单面抛光片、显影液(苏州汶颢芯片科技有限公司);SU-82 075光刻胶(美国Microchem公司)、磷酸盐缓冲盐(PBS)、96培养孔板(美国Corning公司);无菌注射器(江苏恒康公司);PE/20软管(美国Scientific Commodities公司);多聚赖氨酸溶液(北京索莱宝科技有限公司);DMEM/F-12培养基(美国Hyclone公司);胎牛血清(加拿大MULTICELL公司);青霉素-链霉素溶液、胰蛋白酶(美国Gibco公司);CCK-8检测试剂盒(大连美仑生物有限公司);山羊血清、Tween-20溶液(碧云天生物科技公司);荧光素异硫氰酸酯标记的小麦胚芽凝集素(WGA-FITC)、二甲基亚砜(DMSO)、大田软海绵酸、河豚毒素、环亚胺毒素(北京普华仕科技发展有限公司);芋螺毒素(上海楚肽生物科技有限公司);雷公藤甲素(上海陶素生物科技有限公司)。所有化学试剂除特殊说明外均为分析纯。
-
精密称取OA溶解于DMSO,配置浓度为31 μmol/L的储备液;CTX溶解于超纯水中,配置浓度为200 μmol/L的储备液;TTX溶解于0.1%甲酸溶液,配置浓度为313 μmol/L的储备液;5 μmol/L的GYM标准溶液,氮气吹干后培养基复溶,于−20 ℃保存。用培养基稀释至低、中、高三个浓度的溶液:OA(10、50、100 nmol/L)、CTX(0.5、5、20 μmol/L)、TTX(0.04、0.4、4 μmol/L)、GYM(4、10、40 μmol/L)。雷公藤甲素溶解于DMSO,配置浓度为10 mmol/L的储备液,培养基稀释至实验浓度0.5、1、3、10、20、50 μmol/L。
-
人脐静脉内皮细胞(HUVEC)购买自美国ATCC细胞库,培养于含10%胎牛血清和1%青霉素-链霉素的DMEM/F-12培养基中,置于37 ℃、5% CO2培养箱中培养,取对数生长期细胞用于后续实验。
-
使用AutoCAD软件设计、软光刻技术制备三通道并列的微流控芯片,如图1所示。将凝血酶(4 U/ml)、PBS和纤维蛋白原(100 mg/ml)按25∶22∶3比例混匀加入芯片胶原通道;血管通道加入PBS稀释的基质胶(1∶50,V/V)后加入对数生长期的HUVEC细胞(3×106 cells/ml);在外部设备上,使用蠕动泵灌注培养基,人体血流剪切应力为5~20 dyn/cm2,参照剪切应力公式τ=6 μQ/h2w计算流体流速(其中τ为流体剪切应力,μ为流体粘滞度,Q为流体流速,h和w分别为芯片内细胞培养腔的高度和宽度),模拟细胞在体内所受的血流剪切应力,于体外重构心血管微环境[2]。
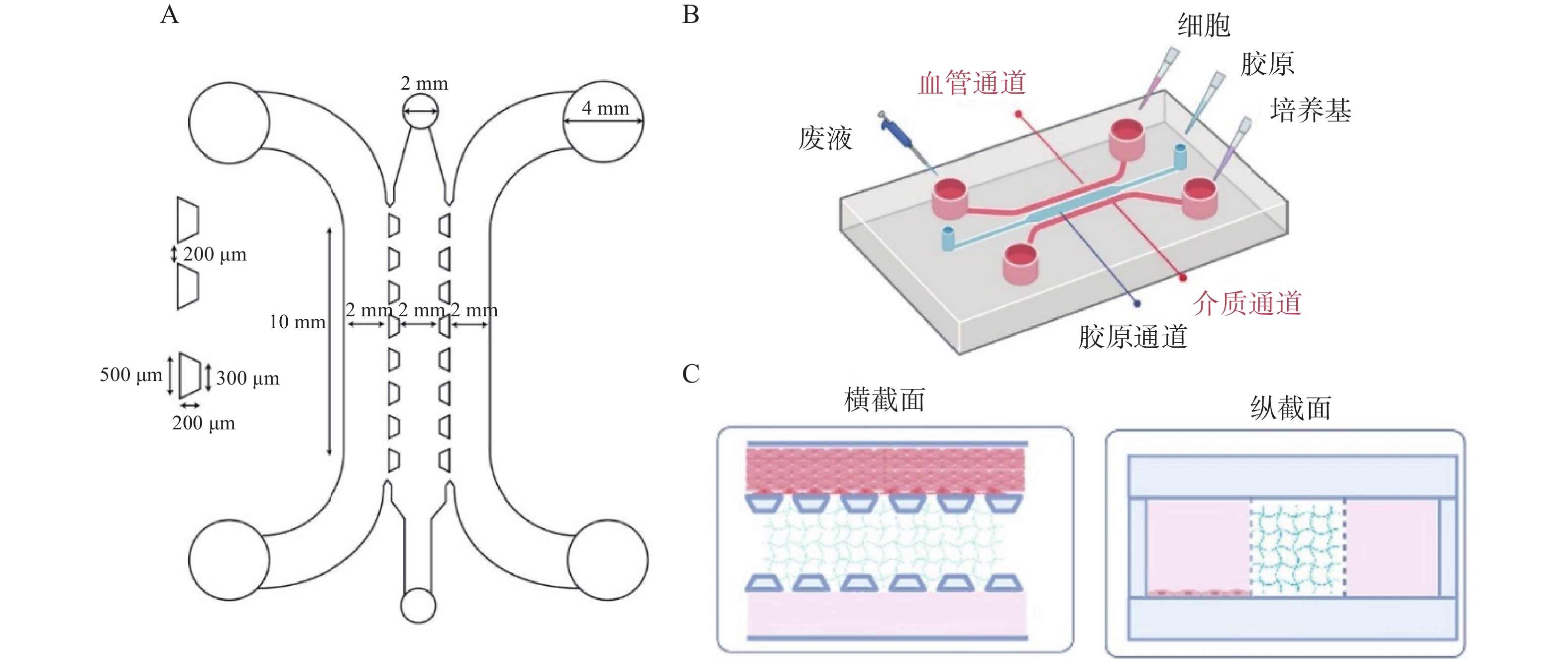
图 1 心血管芯片模拟图
-
取对数生长期的HUVEC细胞,按5 000 cells/孔的细胞密度接种于96孔板,空白组不加细胞。24 h后吸除原培养基,空白组与正常对照组各孔中只加入细胞培养液,实验组中加入含有不同浓度的毒素试液,每孔100 μl,每组6个平行。24 h后观察细胞形态,向各组每孔中加入10%的CCK-8溶液,避光孵育1 h。酶标仪检测450 nm处各样品的吸光度值(OD)计算相对细胞活力。
相对细胞活力(%)=[(OD药物−OD空白)/(OD对照−OD空白)]×100
-
于心血管芯片模型血管通道加入20 μl不同浓度的毒素,24 h后用PBS缓慢润洗通道内细胞。将4%多聚甲醛缓慢注入到芯片微通道内,固定20 min,PBS润洗去除通道内残留的多聚甲醛。加入10%山羊血清的PBST(PBS+0.1% Tween-20),封闭30 min。在通道内加入适量的WGA-FITC(WGA-FITC∶PBS=1∶200,V/V
),避光孵育1 h。芯片通道PBS润洗3次,每次5 min,荧光倒置显微镜下观察并采集数据。根据毒素损伤前后WGA-FITC染色荧光强度(MFI)变化情况计算糖萼损伤率。 糖萼损伤率(%)=[1−(MFI损伤/MFI正常)]×l00
-
考察不同浓度的雷公藤甲素对HUVEC细胞活性的影响,筛选出雷公藤甲素的最佳预保护浓度用于后续实验。取对数生长期的HUVEC细胞接种于96孔板,向实验组、对照组各孔中分别加入雷公藤甲素预保护1 h。实验组加入不同浓度的毒素溶液,CCK-8法评价雷公藤甲素对毒素损伤后HUVEC细胞的保护作用,实验步骤详见“1.5”项。
采用上述构建的心血管芯片模型,于血管通道内加入雷公藤甲素预保护1 h后,实验组加入不同浓度的毒素,考察雷公藤甲素对毒素损伤血管糖萼组织保护作用,实验步骤详见“1.6”项。
-
采用Image J软件计算免疫荧光强度表达,GraphPad Prism 7统计学软件进行数据分析,组间比较采用单因素方差分析,*P<0.05,**P<0.01表示差异有统计学意义。
-
以1 μl/min的流速动态培养细胞,HUVEC细胞在所构建的心血管器官芯片模型中生长状态良好,存活率高。经WGA-FITC免疫荧光染色,可清楚观察到结构完整的血管内皮糖萼,符合体外心血管模型的要求,如图2所示。
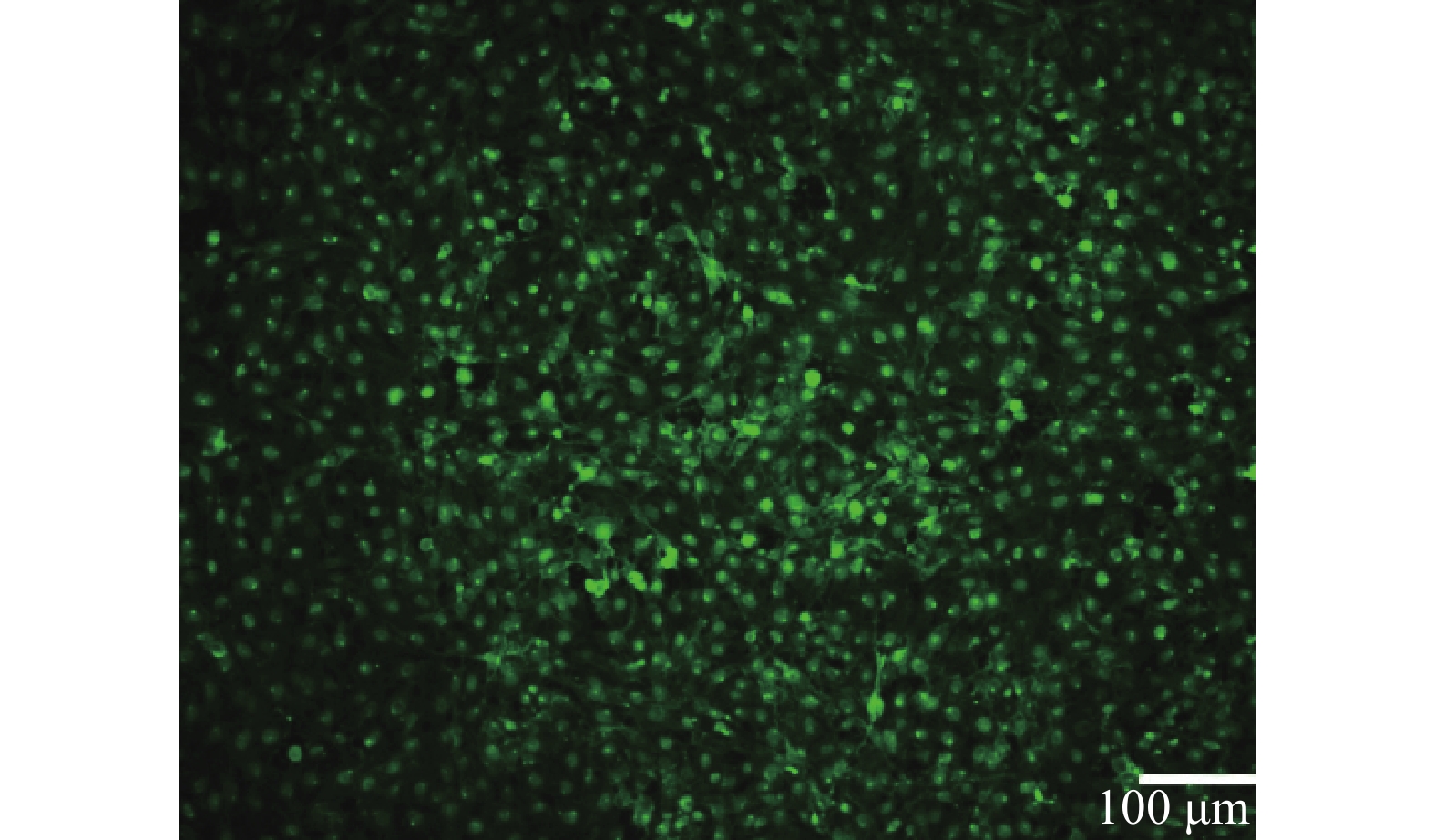
图 2 糖萼免疫荧光染色图
-
采用CCK-8法考察不同浓度的雷公藤甲素对HUVEC细胞活力的影响,结果如图3所示,在测试浓度范围内,雷公藤甲素对HUVEC细胞不存在明显的活力抑制,选择浓度为1 μmol/L的雷公藤甲素进行后续实验。
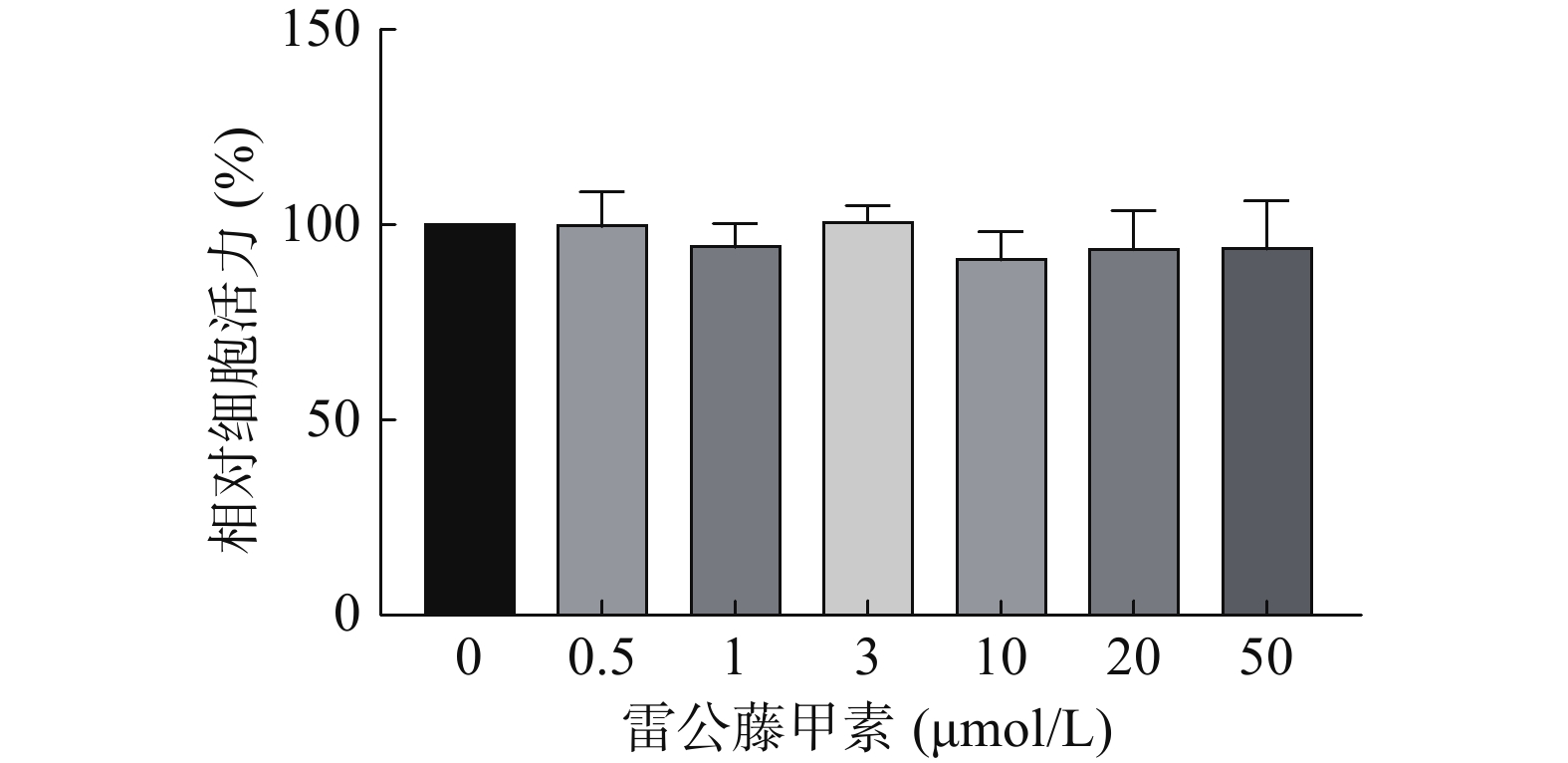
图 3 雷公藤甲素对HUVEC细胞活力评价结果
-
采用CCK-8法测定雷公藤甲素预保护前后,低、中、高三个浓度的毒素对HUVEC细胞活力的影响。与对照组相比,OA在中、高浓度下对细胞活力具有较强的抑制作用(P<0.01),其IC50为54.12 nmol/L。GYM在高浓度对细胞活力有较大影响(P<0.05),但是在加入1 μmol/L雷公藤甲素预保护后,OA与GYM对细胞的毒性显著降低,细胞活力与对照组相比无显著性差异,说明雷公藤甲素对OA和GYM损伤HUVEC细胞具有一定的保护作用。而CTX、TTX在雷公藤甲素预保护前后,对HUVEC细胞活力均无显著抑制作用,结果如图4、5所示。
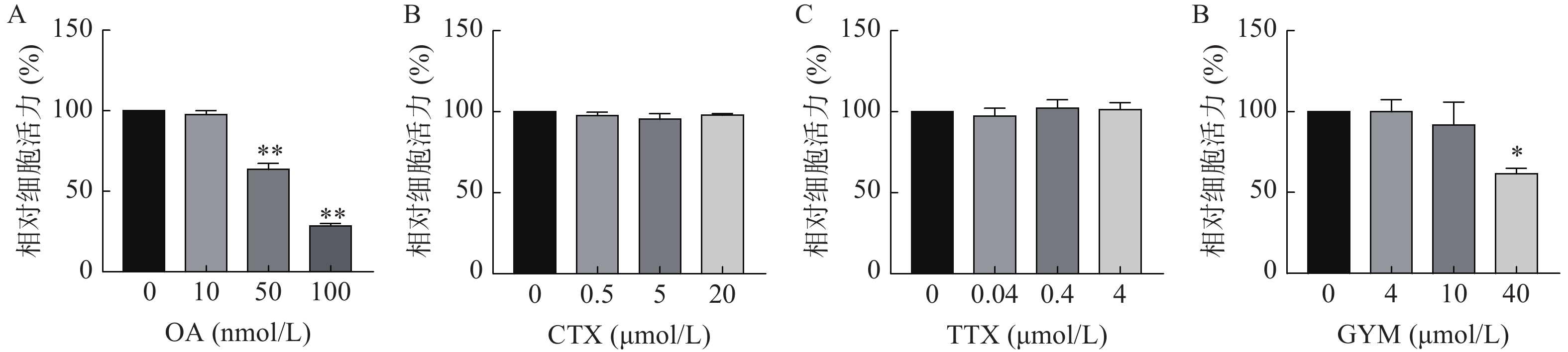
图 4 四种毒素对HUVEC细胞活力评价结果
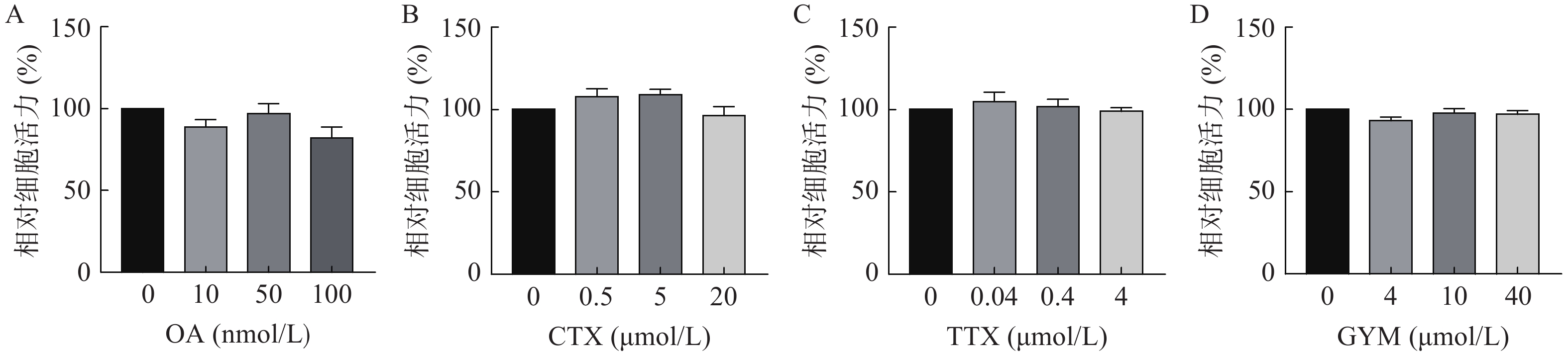
图 5 四种毒素在雷公藤甲素预保护后对HUVEC细胞活力评价结果
-
在所建立的心血管芯片平台上研究雷公藤甲素对四种毒素产生细胞内皮糖萼损伤的保护作用,免疫荧光染色结果如图6和图7所示。OA作用后,HUVEC细胞出现皱缩、变圆、体积变小等形态变化,且随着实验浓度的增大,所呈现的上述细胞形态变化加剧,培养液中可见大量细胞碎片和悬浮的死细胞,说明OA对HUVEC细胞毒性较强,不仅对细胞糖萼有较严重的损伤,还影响了细胞与芯片PDMS基底的贴附水平,心血管芯片结构和功能受损。CTX、TTX和GYM作用后,HUVEC细胞受到不同程度的损伤,内皮细胞糖萼的WGA-FITC染色荧光强度信号与对照组相比均明显降低,具有统计学意义(P<0.01)。经雷公藤甲素预保护后,低浓度OA所产生的细胞损伤得以减轻,糖萼损伤率为28.20%,但是中浓度和高浓度作用下的细胞依旧损伤严重。除OA外,其余三种毒素作用后的糖萼WGA-FITC染色荧光强度均有明显升高,糖萼损伤率见表1。
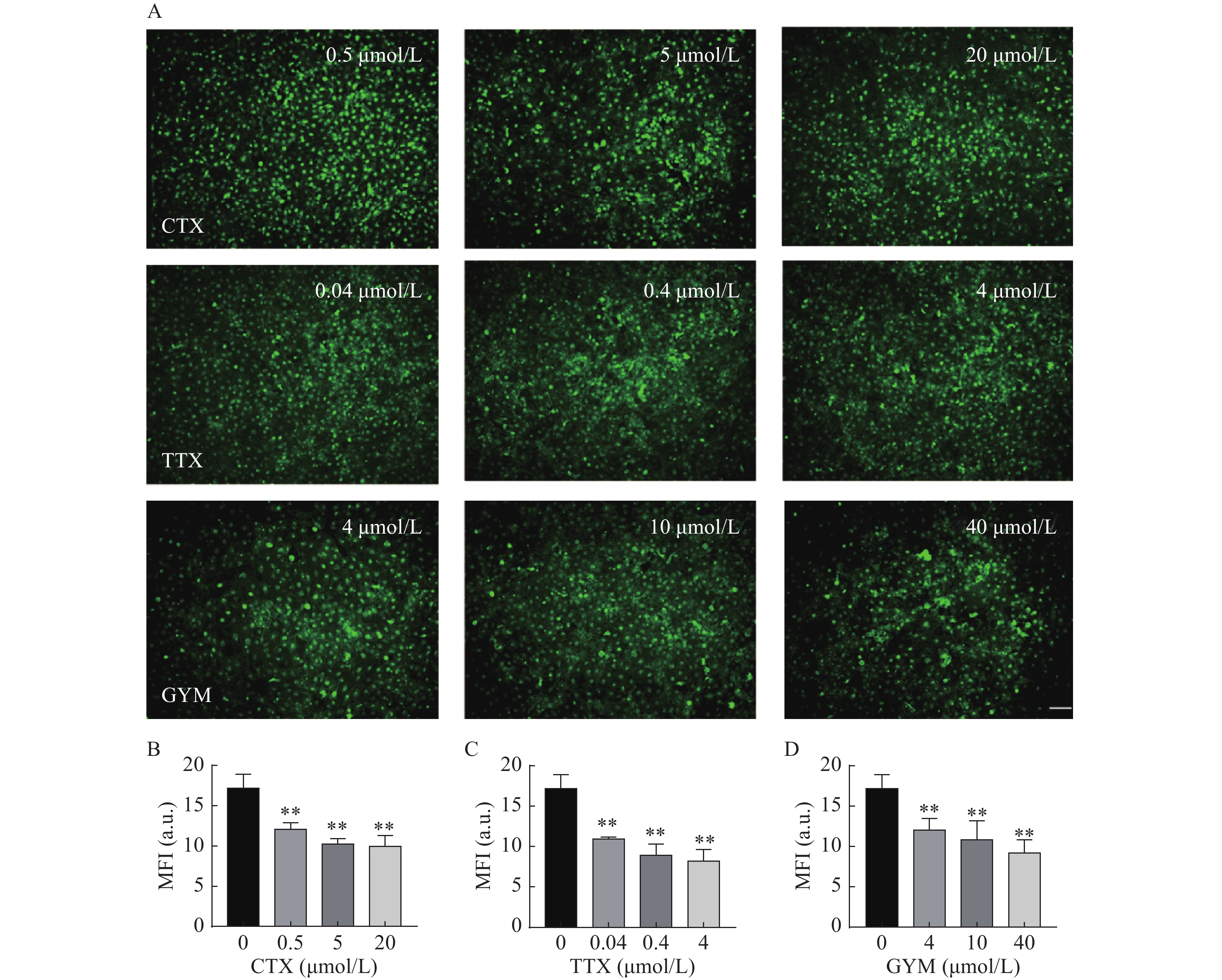
图 6 毒素对心血管芯片上糖萼的损伤情况
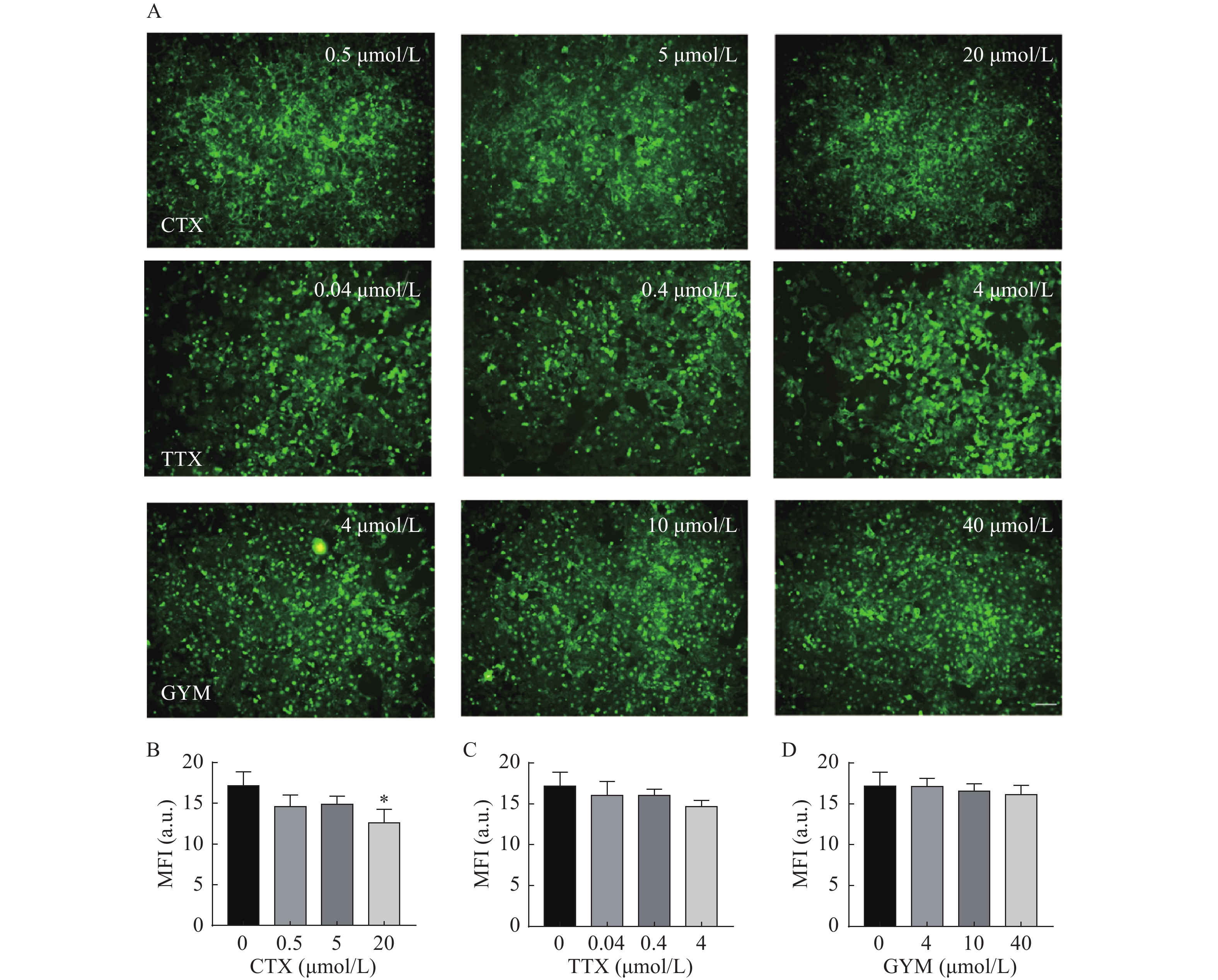
图 7 雷公藤甲素预保护后毒素对心血管芯片上糖萼的损伤情况
表 1 心血管芯片上的糖萼损伤率
毒素 给药浓度(μmol/L) 糖萼损伤率(%) 预保护前 预保护后 CTX 0.5 36.30 14.90 5 48.04 13.47 20 52.28 26.52 TTX 0.04 29.55 6.52 0.4 40.30 6.69 4 41.97 14.60 GYM 4 23.53 0.22 10 25.89 3.65 40 40.41 6.01 -
海洋生物中的活性物质有很多,对海洋生物毒素研究却是冰山一角。海洋生物毒素根据化学结构的不同常分为三类:聚醚类、多肽类和生物碱类毒素[3]。OA是一种聚醚类化合物,是近年来研究最突出的一种海洋生物毒素,研究发现暴露于OA会导致细胞凋亡以及促进肿瘤和癌症的发生,对人体造成很大危害[4]。多肽类毒素以CTX最具代表性,CTX能特异性地作用于神经递质受体以及多种离子通道,不仅可直接作为药物,还可作为分子工具发展新药先导化合物,对神经生物学的研究具有重要意义[5]。TTX是一种生物碱类毒素,可选择性地与钠离子通道受体结合,研究发现,TTX的麻醉效果强,持续时间长,在镇痛、镇静方面应用前景广泛[6]。随着对海洋生物研究的深入,一些新的毒素也陆续被发现。GYM是一种具有环亚胺结构的毒素,相对于其他三种毒素发现较晚、毒性较弱,但其降解速度缓慢,存在长期毒性效应,在生态系统中的潜在风险不容忽视[7]。尽管海洋生物毒素的研究越来越受到人们的重视,但是现有的评价模型限制了毒素研究工作的进一步开展。
微流控器官芯片模型利用微流控技术对流体进行精确调控,模拟生物体内环境,在体外进行器官生理及病理状态的复制,到目前为止,已经实现肝、肾、肠道等多种器官芯片的构建,在药物筛选、药效和毒性评价方面显示出良好的应用前景[8]。大量研究表明,血液循环流动产生的剪切应力对内皮细胞分化、生长状态和功能表达有着重要作用[9]。本研究基于微流控技术构建心血管芯片模型,将HUVEC细胞接种于三通道的微流控芯片中,通过蠕动泵控制培养基流动速度为1 μl/min,有利于营养物质的输送和废物排出,与静态培养相比,细胞所处的动态环境更接近体内生理条件,心血管芯片模型中的细胞生长状态良好,形态呈铺路石样。
糖萼是存在于内皮细胞表面的多糖-蛋白复合物,参与调节炎症反应、血管通透性、凝血平衡在内的多种生理过程,是血管内皮细胞发挥生物功能的重要结构[10]。本研究采用免疫荧光染色法,通过FITC-WGA与HUVEC细胞中糖萼的葡萄糖胺基团选择性结合,测量其荧光强度以反映细胞糖萼损伤情况。结果显示,在实验浓度范围内,OA对细胞具有较大毒性,对糖萼组织损害严重;40 μmol/L的GYM对细胞活力有较强的抑制作用,糖萼损伤率高达40.41%;而TTX和CTX两种毒素对HUVEC细胞无明显活力抑制,但是其糖萼组织受损严重,可见这两种毒素虽然本身对细胞不存在明显的毒性作用,但是通过其他途径与细胞上的结构组织相互作用,从而影响细胞的功能表达。
雷公藤甲素是卫矛科植物雷公藤的主要活性成分,具有抗炎、抗菌、抗肿瘤等药理作用,但是对心血管的作用尚未完全明确。张诗雨等[11]发现雷公藤甲素可作用于血管内皮细胞核转录因子NF-κB,改善脂多糖(LPS)刺激HUVEC细胞所产生的炎症反应。宗佳琪等[12]在LPS诱导的内毒血症大鼠的心血管功能损伤研究中,同样也发现雷公藤甲素对血管内皮相似的保护作用。本研究初步探索了雷公藤甲素对毒素致伤的保护作用,结果显示,经1 μmol/L雷公藤甲素预保护后,四种毒素作用下的细胞活力与对照组相当,对实验浓度范围内CTX、TTX、GYM以及低浓度OA所产生的糖萼损伤具有一定保护作用。
综上,本文构建了一种心血管芯片模拟心血管结构与功能,包括血管表面的糖萼组织。应用该芯片考察了OA、TTX、CTX和GYM四种深海毒素的毒性效应,发现四种毒素对HUVEC细胞活力以及表面的糖萼组织具有损伤,且呈一定的剂量关系,而雷公藤甲素对上述毒素致伤具有保护作用。
Evaluation of toxin-induced injury and protective effect of triptolide based on a cardiovascular chip model
-
摘要:
目的 构建一种心血管芯片模型,评估四种海洋毒素即大田软海绵酸(OA)、芋螺毒素(CTX)、河豚毒素(TTX)和环亚胺毒素(GYM)对血管糖萼组织的损伤,初步探索雷公藤甲素对毒素致伤的保护作用。 方法 将人脐静脉内皮细胞(HUVEC)接种于三通道结构的微流控芯片,并对所构建的心血管芯片模型进行表征。采用CCK-8法和免疫荧光染色法分析低、中、高三个浓度的海洋毒素对细胞活力和糖萼组织损伤情况,以及雷公藤甲素对毒素致伤的保护作用。 结果 所构建心血管器官芯片中的细胞生长状态良好,具有结构完整的糖萼组织,与对照组相比,OA中、高浓度以及GYM高浓度均对细胞活力具有一定抑制(P<0.05),CTX和TTX在所测浓度下对细胞没有明显活力抑制,但是四种毒素均引起细胞表面糖萼组织的严重损伤(P<0.01),且随着浓度增加,糖萼损伤率升高。经雷公藤甲素预保护后,四种毒素对HUVEC的毒性显著降低,糖萼组织损伤率下降。 结论 四种毒素对HUVEC细胞活力以及表面的糖萼组织具有损伤,且呈一定的剂量关系,而雷公藤甲素对毒素损伤后的HUVEC细胞具有保护作用。 Abstract:Objective To construct a cardiovascular chip model for evaluating the damage of vascular glycocalyx induced by four marine toxins: okadaic acid (OA), conotoxin (CTX), tetrodotoxin (TTX) and gymnodimine (GYM), and explore the protective effect of triptolide on toxin-induced injury. Methods Human umbilical vein endothelial cells(HUVEC) were inoculated into a three-channel microfluidic chip. CCK-8 method and immunofluorescence staining were used to analyze the damage of cell viability and glycocalyx tissue induced by low, middle and high concentrations of marine toxin, as well as the protective effect of triptolide on toxin-induced injury. Results The cells in the cardiovascular chip grew well and had structurally intact glycocalyx. Compared with the control group, the activity of HUVEC cells were inhibited in group of the medium and high concentration of OA and high concentration of GYM (P<0.05). The activity of cells had not been inhibited by CTX and TTX significantly , but all the four toxins caused serious damage to the glycocalyx tissue (P<0.01). After pre-protection with triptolide, the toxicity of the four toxins to HUVEC cells and the damage rate of glycocalyx decreased significantly. Conclusion The four marine biotoxins could damage the activity and glycocalyx of HUVEC cells in a dose-dependent manner, while triptolide has a protective effect on HUVEC cells injured by toxin. -
Key words:
- cardiovascular chip /
- marine biotoxins /
- glycocalyx damage /
- triptolide
-
骨骼肌约占人体重的35%,是人体内最大的器官系统之一,为人体提供运动所必须的产能,储能以及帮助调节体温[1]。骨骼肌具有强大的再生能力,这对肌肉损伤后修复及其他导致肌病具有重要意义[2]。已经有许多研究报道非肌源性细胞在再生过程中起到关键作用,比如以髓系为代表的免疫细胞被募集到损伤部位对肌肉修复发生产生影响[3]。最近的文献进一步揭示髓系细胞中单核细胞及巨噬细胞在骨骼肌再生反应中起到关键作用[4]。
随着流式分析仪器和试剂的进步,多色流式实验目前已被广泛应用[5]。多色流式细胞术的优势在于能够同时使用多个标记物进行单个细胞层面研究,分析组织中不同细胞群之间的关联性,同时也可以减少样本用量,提高样本检测通量,提高实验效率[6]。特别是在免疫细胞的研究方面,多色流式可以最大程度的同时检测样本中的不同免疫细胞表面蛋白表达程度,并直观的以降维分析方式展现出来,具有广阔的应用前景[7]。
本研究采用流式细胞仪对小鼠损伤腓肠肌的单细胞悬液进行数据采集。同时应用t-分部邻域嵌入算法(t-SNE)[8]和手动圈门分析所得结果,为流式数据分析和结果展示提供一种新的方法和思路。
1. 材料和方法
1.1 实验动物
雄性C57BL/6J(B6)同系小鼠用于小鼠骨骼肌流式分析实验。实验小鼠来自中国同济大学实验动物中心。所有动物实验遵守动物伦理指南,已获海军军医大学药物实验伦理委员会批准。
1.2 实验试剂
流式抗体为FITC标记的CD11b、APC-A700标记的F4/80、APC-A750标记的MHC II、PB450标记的Ly6G,BV610标记的CD45、BV650标记的CD11c、PE标记的Ly6C,抗体均购自美国Biolegend公司。Ⅱ型胶原酶购自瑞士Roche,心脏毒素购自美国MCE。
1.3 仪器与设配
流式细胞仪(Beckman CytoFLEX S,美国),离心机(湘仪,中国)。
2. 方法
2.1 小鼠肌肉损伤模型
使用1%的戊巴比妥钠麻醉小鼠,假手术组对小鼠腓肠肌注射50 μl生理盐水,模型组对小鼠腓肠肌注射50 μl 10 μmol/L的心脏毒素,均在损伤7 d后处死小鼠。
2.2 小鼠腓肠肌单细胞悬液制备
颈椎脱臼法处死小鼠,使用酒精棉球擦拭消毒小鼠腿部表面后解剖小鼠,从两条腿上分离出腓肠肌。将其放入20 mm培养皿中,加入5 ml提前预冷的磷酸盐缓冲液(PBS)。使用锋利的剪刀将腓肠肌剪碎,加入浓度为5 mg/ml的Ⅱ型胶原酶5 ml,置于37 ℃孵箱孵育60 min。孵育后将细胞悬液通过40 μm尼龙筛网过滤,转移至新的15 ml离心管中。4 ℃,800×g离心5 min,弃掉上清液,将所得沉淀转移至5 ml流式管中,加入100 μl PBS重悬,即为腓肠肌单细胞悬液。
2.3 流式细胞染色及上机检测
用PBS将细胞浓度调节至每100 μl约1×107个细胞。每100 μl液体中0.5 μl流式抗体,4 ℃孵育45 min。孵育后4 ℃,800×g离心5 min。弃掉上清液,用200 μl PBS重悬细胞,上机检测。
2.4 分析及统计方法
流式数据应用FlowJoTM v10.8.1流式分析软件(购自美国BD公司)对小鼠腓肠肌细胞进行数据分析。数据分析与图形呈现通过GraphPad Prism软件执行。实验数据主要以平均值±标准误(Mean±SEM)形式展示,两个独立样本之间采用t检验(t-test)进行两组样本的比较。P<0.05表示有显著性差异。
3. 结果
3.1 小鼠腓肠肌流式细胞分析结果
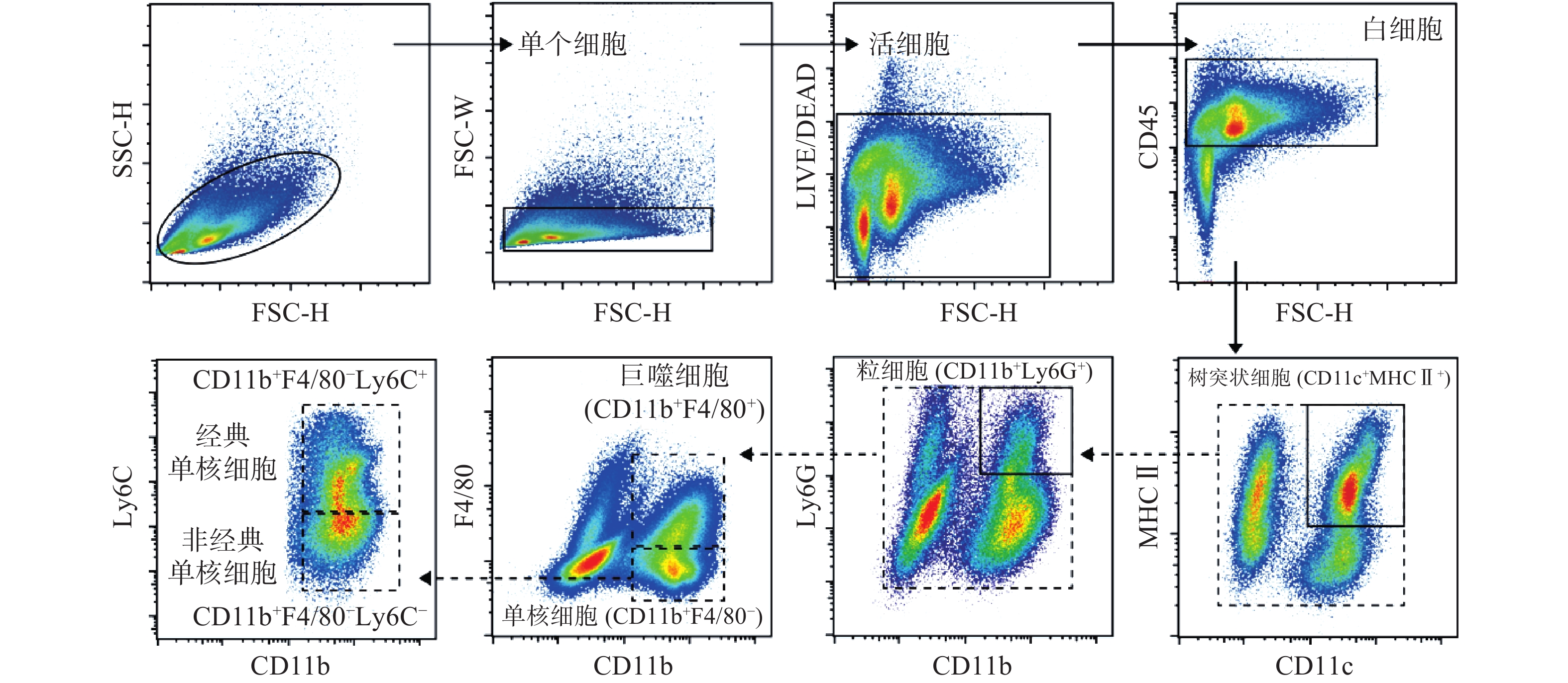
使用流式细胞术检测小鼠腓肠肌单细胞悬液中单核细胞圈门策略如图1所示。首先通过FSC-H和SSC-H圈出主要细胞群,根据FSC-H与FSC-W去除粘连细胞,通过LIVE/DEAD圈出活细胞。CD45用于圈选出白细胞群,结合CD11c、MHCⅡ、CD11b、F4/80、Ly6C和Ly6G圈选出CD11c阳性与MHCⅡ阳性树突状细胞、CD11b阳性与Ly6G阳性粒细胞、CD11b阳性与F4/80阳性巨噬细胞,Ly6C阳性单核细胞。各散点图中细胞分群界限清晰,流式圈门设置合理,为后续进一步的 t-SNE分析与手动圈门分析提供良好的数据基础。
3.2 小鼠腓肠肌细胞t-SNE分析
将经流式细胞仪采集的小鼠腓肠肌免疫细胞(CD45阳性)数据进行降维分析。如图2所示,每一个点对应一个细胞,图2B中根据荧光通道表达强弱进行着色,分别表征CD11b、CD11c、Ly6G和Ly6C的表达水平。t-SNE降维分析将高维的细胞数据以二维平面的方式直观呈现出来,这种形式不同于以往流式分析中的逐级圈门方式,能将各群细胞的抗原表达情况更加准确全面,直观地展现出来[8]。通过图2A可以看出假手术组与模型组的腓肠肌中免疫细胞构成发生了明显变化,假手术组主要以黑色圆圈1标志的细胞群为主,而在模型组中,主要以红色圈2、3、4、5标志的细胞群为主。结合图2B中CD11b、CD11c、Ly6C和Ly6G表达水平图,可以发现红色圆圈3、4、5细胞群高表达Ly6C,揭示造模后的免疫细胞浸润主要以高表达Ly6C的阳性细胞为主,同时模型组中的CD11b、CD11c、Ly6G表达水平明显高于假手术组,提示模型组炎症水平高于假手术组。总体来看,t-SNE分析结果直观明了地反映了细胞群体之间的相互关系和整体情况,为下一步手动圈门分析提供了指引方向。
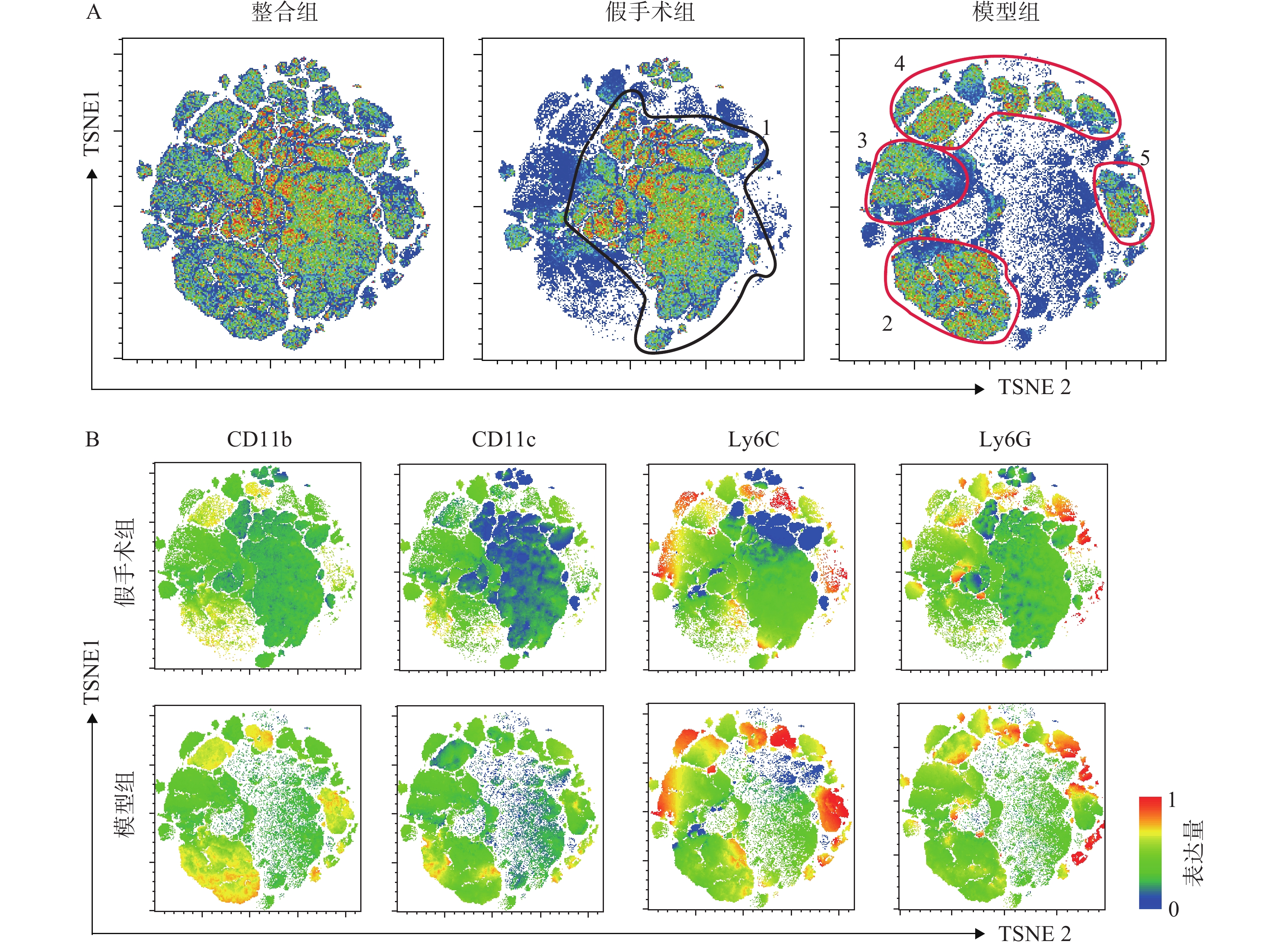 图 2 小鼠腓肠肌单细胞悬液单核细胞降维图A. 整合组(假手术组+模型组)、假手术组、模型组t-SNE降维结果;B. 假手术组与模型组CD11b、CD11c、Ly6C和Ly6G表达水平
图 2 小鼠腓肠肌单细胞悬液单核细胞降维图A. 整合组(假手术组+模型组)、假手术组、模型组t-SNE降维结果;B. 假手术组与模型组CD11b、CD11c、Ly6C和Ly6G表达水平3.3 小鼠腓肠肌细胞手动圈门分析
对小鼠CD45阳性免疫细胞进行进一步手动圈门分析,得到各细胞类型具体占比。如图3所示,从CD11c阳性细胞群中手动圈门发现模型组中MHCⅡ阳性树突状细胞比例显著高于假手术组(图3A);与假手术组相比,模型组中Ly6G阳性粒细胞占CD11b阳性细胞比例显著增加(图3B);F4/80阳性巨噬细胞占CD11b阳性细胞在模型组中显著增加,与前期的tSNE结果一致。值得注意的是,与假手术组相比,在CD11b阳性F4/80阴性细胞群中手动圈出的Ly6C阳性单核细胞群与树突状细胞、粒细胞和巨噬细胞相比,在模型组中增加比例最为明显。以上结果表明在肌肉损伤模型7 d后的模型中以树突状细胞、粒细胞、单核细胞、巨噬细胞为代表的髓系细胞免疫细胞浸润依然明显,并且在这之中单核细胞增加比例最多。
 图 3 小鼠腓肠肌单细胞悬液髓系细胞手动圈门图(n=6)A. 假手术组与模型组中树突状细胞比例;B. 假手术组与模型组中粒细胞细胞比例;C. 假手术组与模型组中巨噬细胞比例;D. 假手术组与模型组中单核细胞比例**P<0.01,***P<0.001,与假手术组比较。
图 3 小鼠腓肠肌单细胞悬液髓系细胞手动圈门图(n=6)A. 假手术组与模型组中树突状细胞比例;B. 假手术组与模型组中粒细胞细胞比例;C. 假手术组与模型组中巨噬细胞比例;D. 假手术组与模型组中单核细胞比例**P<0.01,***P<0.001,与假手术组比较。4. 讨论
骨骼肌对很多生理病理造成的损伤有强大的修复能力[1],肌肉损伤修复是一个需要多类细胞协同合作的复杂过程[9-12],其中非肌源性细胞,特别是免疫细胞对损伤过程修复的干预有着十分重要的意义[10]。本文主要用流式细胞仪对小鼠骨骼肌部分髓系细胞进行分析,为后续的流式分选及单细胞测序实验指引了方向。
随着流式分析试剂和仪器的进步,多色流式实验已经广泛应用于各项研究,同传统2~3色的流式实验相比,多色流式能够同时使用多个标记物对单个细胞进行研究,分析多个细胞群之间的关联性,更全面地展示整体免疫微环境的变化。流式细胞数据分析常见软件包括Flowjo、FACS Diva、CytoBank等,可以为诸如细胞表型、荧光蛋白、细胞周期、细胞凋亡、细胞增殖、磷酸化蛋白检测、活性氧检测等提供便捷专业的分析。其中,FlowJo是较早为流式分析设计的软件,有多类插件可供选择,能将数据以csv文件导出,给后续的生物信息学分析提供了极大的便利。降维分析相较于传统的逐级圈门方法,减少了人为圈门引入的误差,通过整体分析也更有助于发现新的细胞群体和获得更为直观的整体表达情况。目前随着流式细胞术的不断深入发展,流式数据分析方法也不断更新,结合降维分析的方法为多色实验和单细胞测序实验都奠定了一定的基础。然而,降维方法本身并不能确认结果的正确与否,同时引入手动圈门也可能引入更大的人为误差。t-SNE虽然能够将高维数据以二维形式展示,但在降维过程中不可避免地会丢失部分信息,可能导致某些细胞群体的特征被忽略。除此之外,降维结果十分依赖于参数的设置,不同参数设置往往会导致结果差异巨大。因此,在使用这些方法时一定要根据数据本身的特性进行合适的参数选择,确保保留并尽可能多地提取到数据中有效信息。操作过程中,研究人员应通过多次实验和参数调优来找到最佳设置,以便最大限度地减少信息丢失和偏差。同时,结合其他分析方法,如统一流形逼近与投影法(UMAP)或主成分分析(PCA)等,通过对比不同降维方法的结果,进一步提高数据的可靠性和全面性。这不仅能提高对相关生物过程的理解,还能为未来的研究提供更为坚实的基础。
T-SNE能够将高维数据有效地降维成二维展示,使得复杂的细胞群体关系变得更加直观和易于理解。这种方法特别适用于大规模细胞数据的分析,如免疫细胞群体的分类和功能研究。在目前的研究中,t-SNE降维结合手动圈门的方法不仅提高了细胞群体的分辨率,还能更精确地捕捉到细胞间的微小差异。这对于理解骨骼肌损伤修复过程中不同髓系细胞的角色和动态变化具有重要意义。未来,这种方法可以进一步应用于其他生物医学领域,如肿瘤微环境分析、自身免疫疾病研究以及再生医学等。此外,随着机器学习和人工智能技术的发展,t-SNE等降维方法有望与这些技术结合,实现对流式细胞数据的自动化和智能化分析。这不仅能提高数据处理的效率,还能减少人为误差,提供更加客观和可靠的分析结果。本研究关注的小鼠髓系细胞是肌肉损伤修复的重要基础,但缺乏其他非肌源性细胞比如纤维成脂祖细胞(FAPs)、淋巴细胞、血管内皮细胞(VEC)等信息,展示肌肉损伤的微环境还不够全面。在未来的研究中,将把以上的细胞群体一并纳入分析,基于流式细胞术与生物信息学结合,推动研究更往深层次发展。
总的来说,自动降维分析可以将高维数据转换为低维空间,从而直观地对细胞进行聚类和分群。与此同时,手动圈门分析提供了对特定细胞群的精细控制和确认,可以根据已知的生物学标志物或功能特性对细胞进行分类和验证。这种方法的灵活性和直观性有助于深入理解细胞群的特征和功能。通过结合这两种分析方法,我们不仅可以获得全局性的细胞分布和关系图景,还能在具体的细胞群层面上进行精细化分析,能够提高数据解读的准确性和深度,从而更好地揭示生物学研究中的复杂现象,具有深远的科研意义。
-
图 7 雷公藤甲素预保护后毒素对心血管芯片上糖萼的损伤情况
A.糖萼免疫荧光染色图; B~D.分别为芋螺毒素、河豚毒素、环亚胺毒素的糖萼量化分析结果,*P<0.05,与对照组比较
表 1 心血管芯片上的糖萼损伤率
毒素 给药浓度(μmol/L) 糖萼损伤率(%) 预保护前 预保护后 CTX 0.5 36.30 14.90 5 48.04 13.47 20 52.28 26.52 TTX 0.04 29.55 6.52 0.4 40.30 6.69 4 41.97 14.60 GYM 4 23.53 0.22 10 25.89 3.65 40 40.41 6.01  下载: 导出CSV
下载: 导出CSV
-
[1] BARRECA M, SPANÒ V, MONTALBANO A, et al. Marine anticancer agents: an overview with a particular focus on their chemical classes[J]. Mar Drugs,2020,18(12):619. doi: 10.3390/md18120619 [2] YOSHIZUMI M, ABE J I, TSUCHIYA K, et al. Stress and vascular responses: atheroprotective effect of laminar fluid shear stress in endothelial cells: possible role of mitogen-activated protein kinases[J]. J Pharmacol Sci,2003,91(3):172-176. doi: 10.1254/jphs.91.172 [3] 陈巧莉, 杨兵, 洪晴悦, 等. 海洋生物毒素的分类、毒害作用机制及检测技术研究进展[J]. 食品科学, 2021, 42(5):321-331. doi: 10.7506/spkx1002-6630-20200326-380 [4] FU L L, ZHAO X Y, JI L D, et al. Okadaic acid (OA): Toxicity, detection and detoxification[J]. Toxicon,2019,160:1-7. doi: 10.1016/j.toxicon.2018.12.007 [5] JIN A H, MUTTENTHALER M, DUTERTRE S, et al. Conotoxins: chemistry and biology[J]. Chem Rev,2019,119(21):11510-11549. doi: 10.1021/acs.chemrev.9b00207 [6] BUCCIARELLI G M, LECHNER M, FONTES A, et al. From poison to promise: the evolution of tetrodotoxin and its potential as a therapeutic[J]. Toxins,2021,13(8):517. doi: 10.3390/toxins13080517 [7] OTERO P, SILVA M. Emerging marine biotoxins in European waters: potential risks and analytical challenges[J]. Mar Drugs,2022,20(3):199. doi: 10.3390/md20030199 [8] KIMURA H, SAKAI Y, FUJII T. Organ/body-on-a-chip based on microfluidic technology for drug discovery[J]. Drug Metab Pharmacokinet,2018,33(1):43-48. doi: 10.1016/j.dmpk.2017.11.003 [9] CUCULLO L, HOSSAIN M, PUVENNA V, et al. The role of shear stress in Blood-Brain Barrier endothelial physiology[J]. BMC Neurosci,2011,12:40. doi: 10.1186/1471-2202-12-40 [10] TARBELL J M, CANCEL L M. The glycocalyx and its significance in human medicine[J]. J Intern Med,2016,280(1):97-113. doi: 10.1111/joim.12465 [11] 张诗雨, 张静, 高攀, 等. 雷公藤甲素对血管内皮细胞炎症反应的影响及机制研究[J]. 时珍国医国药, 2021, 32(7):1537-1541. doi: 10.3969/j.issn.1008-0805.2021.07.01 [12] 宗佳琪, 王烁阳, 苏萍, 等. TLR4/NF-κB p65信号通路介导雷公藤甲素对内毒血症大鼠内皮保护作用[J]. 中国中药杂志, 2019, 44(22):4912-4917. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3528
- HTML全文浏览量: 1321
- PDF下载量: 20
- 被引次数: 0
