

 下载:
下载:
 下载:
下载: 

</td></tr></tbody>
</table></div></foreignObject></svg>)
-
外泌体(exosome)是细胞产生的胞外囊泡的一种,在20世纪80年代末首次被描述[1-2]。外泌体作为一种细胞间近距离通讯的特殊介质,在包括免疫反应、抗原递呈和信号转导[3]等各种生理过程中发挥着重要作用。几乎所有真核细胞都可以分泌外泌体,包括脂肪细胞、上皮细胞、成纤维细胞、神经元、星形胶质细胞等,外泌体几乎存在于所有体液如脑脊液、尿液、精子、唾液、血液、玻璃体和乳汁等[4],含有丰富的核酸、蛋白、脂质和代谢物等。外泌体所携带的物质由于源细胞的类型及其所处状态(例如转化、分化、刺激和压力)不同而存在很大差异,由其大小、内容物含量、对受体细胞的功能影响以及细胞来源来概念化,因此是一个高度异质性的群体,具有独特的诱导生物学反应的能力,也为一些疾病如代谢性疾病、心血管疾病、神经退行性疾病、肿瘤等提供诊断和预后信息[5]。此外,作为一种纳米粒径的内源性囊泡,高生物相容性、天然的归巢性能、以及进行功能化修饰后实现更特异性的组织器官和病灶部位的富集,都使得外泌体作为一种新型药物递送载体具有很好的研究价值。本文就外泌体的提取分离方法,在疾病中的诊疗作用以及工程化修饰后用于药物递送做一综述。
-
外泌体的发生机制主要涉及质膜的双重内陷。质膜的第一次内陷形成早期分选内含体(ESEs),此时ESEs膜内表面上附着细胞表面蛋白和细胞外环境相关的可溶性蛋白,有时新形成的ESEs可直接与预先存在的ESEs合并。ESEs在高尔基体和内质网的帮助下形成晚期分选内含体(LSEs)并再次内陷形成多泡体(MVBs),MVBs包含数个腔内小泡(ILVs)。MVBs可以与溶酶体或自噬体融合以被降解,也可以与质膜融合以释放包含的ILVs,ILVs释放后即形成直径约40~160 nm的外泌体[6-7]。
目前常用的外泌体分离方法有超速离心法、密度梯度离心法、超滤法、色谱法、免疫亲和法、聚合物沉淀法等,分别采用不同的分离机制,所得到的外泌体具有不同的得率和纯度,其主要原理和优缺点见表1。
表 1 外泌体的分离方法及特点
方法 原理 优势 耗时 纯度 产率 不足 超速离心法[8] 大小和密度不同的组分具有不同的沉积速度 金标准,适用于大批量样品,技术成熟 >4 h 中 低 仪器昂贵、操作繁琐耗时、产量低,可能会破坏外泌体[9] 密度梯度离心法 大小和密度不同的组分具有不同的沉积速度 高纯度,避免外泌体损伤 >16 h 高 低 前期准备、操作繁琐、耗时[10] 超滤法[11] 不同粒子粒径和相对分子质量的差异 操作简便,不需要特殊设备和试剂 <4 h 高 中 滤膜易堵塞,小粒径外泌体
易丢失[12]色谱法 不同粒子粒径和相对分子质量的差异 简单、经济,能较好保持外泌体生物功能和结构[13] <0.3 h 高 高 需要特殊的柱子和填料,存在脂蛋白污染 免疫亲和法[14] 抗体与外泌体特异性膜蛋白的相互作用 特异性分离外泌体 4~20 h 高 中 昂贵,耗时,分离效果取决于抗体的特异性 聚合物沉淀法[15] 外泌体在高亲水性聚合物影响下溶解度或分散性的变化 操作简单,适用于大体积
样品0.3~12 h 低 高 潜在污染物(提纯蛋白质聚集体或残留聚合物) -
外泌体与免疫和炎症反应相关疾病、心血管疾病、神经系统疾病以及肿瘤等的发生发展有关,递送至受体细胞的蛋白质、代谢物和核酸有效地改变了它们的生物反应,这种外泌体介导的反应可以促进或抑制疾病的进程。作为一种通讯介质,外泌体具有调节复杂细胞内通路的特性。作为一种治疗工具,外泌体避免了如不受控制的细胞分裂、致瘤性以及血管栓塞等与细胞治疗相关的安全问题[16]。除了治疗因素,外泌体具有辅助疾病诊断的特性,它们存在于几乎所有生物体液中,通过对生物体液取样很容易获得并检测其内容物组成,从而对疾病进展做出判断以及确定治疗方法。
-
一些外泌体中的核酸物质可以作为疾病诊断的特异性分子。Niu等[17]发现血清外泌体中的miR-155与肝硬化的进展和肝硬化的临床预后指标密切相关,表明富含miR-155的外泌体可作为肝纤维化诊断和进展的非侵入性生物标志物。在CCl4诱导的肝纤维化小鼠模型中,脂多糖处理的巨噬细胞外泌体高表达miR-500能通过抑制MFN2促进肝星形细胞活化和肝纤维化。因此,血清外泌体的miR-500也可作为肝纤维化的生物标志物[18]。Rong等[19]的研究表明,在SD大鼠肝纤维化模型中,骨髓间充质干细胞外泌体(BMSC-Exo)通过抑制 WNT/β-catenin通路降低了CCl4诱导肝纤维化的能力,能够有效减轻肝纤维化,包括减少胶原蛋白积聚,增强肝功能,抑制炎症和增加肝细胞再生。类似地,Damania等[20]的研究发现,在HepG2细胞的2D/3D培养条件的体外肝损伤模型以及由CCl4引起的大鼠急性肝损伤模型中,大鼠BMSC-Exo具有抗凋亡、抗氧化和促细胞存活作用。能明显减少缺血再灌注损伤,可用于临床肝移植或治疗急性肝功能衰竭。
-
源自骨关节炎软骨细胞的外泌体可以通过miR-449a刺激炎症小体活化并增加巨噬细胞中成熟的IL-1β产生,可能加重滑膜炎并加速骨关节炎的进展[21]。BMSC-Exo同时具有免疫调节作用和抗炎作用,Casado等[22]在抗原触发的猪滑膜炎模型中发现,BMSC-Exo处理后,模型动物关节中的白细胞数量没有明显变化,但淋巴细胞显著减少,并伴随着滑液中的炎性细胞因子显著下降,对治疗滑膜炎具有积极意义。Qin等[23]测试了BMSC-Exo在体外调节成骨细胞活性和体内骨再生中的作用,与miR-27a和miR-206处理相比,用miR-196a处理的成骨细胞表现出最好的成骨活性。miR-196a是典型的成骨相关miRNA,在BMSC-Exo中高度富集,证明了BMSC-Exo在调节成骨细胞分化和成骨基因表达中的重要作用。Chen等[24]发现软骨细胞的外泌体可以在皮下环境中诱导软骨祖细胞构建体的有效异位软骨形成,这可能代表了一种用于软骨再生的无细胞治疗方法。
-
动脉粥样硬化是最常见的心血管疾病之一。Hergenreider[25]发现人脐静脉内皮细胞在剪切力刺激下分泌的外泌体富含miR-143/145,其控制共培养平滑肌细胞中靶基因KLF2的表达,从而减少ApoE(-/-)小鼠主动脉粥样硬化的形成。冠状动脉粥样硬化斑块破裂或侵袭后,易产生急性冠状动脉综合征(ACS)。外泌体中的丰富miRNA可以成为诊断和治疗ACS的重要工具[26]。有研究发现ACS患者血清中的外泌体miR-208a水平显著高于健康个体[27]。心肌缺血再灌注损伤是指心肌组织供血中断一段时间后恢复,但心肌组织损伤加重的现象。Lai等[28]在小鼠急性心肌缺血再灌注损伤的心脏模型中发现,间充质干细胞通过减少心肌梗死面积产生心脏保护作用,而其保护作用主要通过旁分泌中的外泌体起效。Zou等[29]以H9C2细胞系建立大鼠心肌缺血再灌注模型用于研究心肌发病过程,BMSC-Exo处理后的缺血再灌注过程中对细胞增殖、迁移以及心肌细胞凋亡的抑制作用,表现为Apaf1(凋亡蛋白酶激活因子1)表达被显著抑制,ATG13(自噬相关蛋白13)表达在体内显著增加,证明BMSC-Exo还可以通过调节自噬机制抑制与心肌梗死相关的心肌损伤。心力衰竭由多种因素带来的心肌收缩功能障碍引起,有研究者在小鼠压力超负荷诱导的心脏肥大模型中发现小鼠BMSC-Exo通过抑制心肌病理性肥大、抑制心肌细胞凋亡和减少心脏纤维化,在抑制心肌重构中产生重要作用,为心力衰竭等疾病提供了全新有效的治疗策略[30]。
-
基于亲代细胞,外泌体可用作神经退行性疾病、脑创伤、卒中等中枢疾病诊断预后的潜在生物标志物,或产生治疗作用。如人脐静脉内皮细胞衍生的外泌体可以通过传递miR-21-3p抑制ATG12信号传导来减弱缺氧/复氧诱导的神经元细胞凋亡[31]。在Wistar大鼠中风模型中,BMSC-Exo处理的动物组皮层和纹状体的缺血边界区轴突密度和突触素阳性区域增加,增强了神经突重塑、神经发生和血管生成,从而实现中风后脑功能恢复[32]。Zhang等[33]在Wistar大鼠受控皮质冲击诱发的创伤性脑损伤(TBI)模型中使用人BMSC-Exo治疗TBI,发现显著增加了病变边界区和齿状回新生内皮细胞的数量,增加了齿状回新生成熟神经元的数量,并减少了神经炎症,同样证明了BMSC-Exo对脑功能恢复和神经血管重塑的促进作用。在神经退行性疾病中,BMSC-Exo可以再生神经并改善神经退行性疾病的认知功能和记忆障碍[34]。Cui等[35]发现经缺氧预处理的间充质干细胞外泌体抑制了β淀粉样蛋白的积累,并增强了转基因APP/PS1阿尔茨海默病模型小鼠大脑中的突触蛋白表达,此外,星形胶质细胞和小胶质细胞的活化减少、炎症因子如抗炎细胞因子IL-4、IL-10增加和促炎细胞因子TNFα和IL-1β的减少得到验证。作为诊断标记物,脑脊液外泌体中的miRNA包括let-7f-5p、miR-27a-3p、miR-125a-5p、miR-151a-3p和miR-423-5p可作为生物标志物用于帕金森病的早期诊断,从分子水平发挥疾病诊断的精确作用。Kojima等[36]发现负载过氧化氢酶mRNA的外泌体能够减轻6-羟基多巴胺(6-OHDA)或LPS诱导的PD小鼠模型中的神经毒性和神经炎症。加载过氧化氢酶的外泌体在体外PC12神经元细胞和PD的体内6-OHDA模型中引发神经保护作用[37]。
-
外泌体携带的DNA、RNA、蛋白质和代谢产物通过自分泌和旁分泌影响受体细胞,从而在肿瘤的发生、发展、免疫、耐药等方面起到重要作用。多发性骨髓瘤患者的骨髓间充质干细胞外泌体含有较高的miRNA-15a及较多的细胞因子和黏附分子,能够促进多发性骨髓瘤细胞增殖[38]。乳腺癌细胞外泌体含有的miR-105降低了ZO1在内皮细胞中的表达,破坏了内皮细胞的紧密连接,从而增强肿瘤细胞的血管内渗透[39]。肿瘤衍生的外泌体或胞外囊泡可激活免疫反应,外泌体将肿瘤抗原热休克蛋白HSP70-80和MHC-I分子转移到DC细胞,从而对小鼠肿瘤产生有效的CD8+T细胞依赖性抗肿瘤作用[40- 41]。巨噬细胞外泌体通过转运miR-365激活了胰腺癌细胞的胞苷脱氨酶,降低其对吉西他滨的敏感性,显示出外泌体在肿瘤耐药性中的作用[42]。外泌体还可以作为肿瘤诊断和预后的生物标志物。例如对循环外泌体DNA检测能够发现KRASG12D和TP53R273H突变,它们是胰腺癌的潜在生物标志物[43]。血浆外泌体的PD-L1水平与头颈癌的疾病进展有关[44-45],还能预测黑色素瘤患者对PD1抗体治疗的反应[46]。
-
外泌体是一种小粒径的内源性微粒,具有靶向组织特异性、体循环稳定性、良好的生物相容性等优点。通过不同的载药方式,外泌体可以负载不同的药物,通过与源细胞特殊的亲和作用、归巢效应等自然趋向作用,将负载的药物运送到靶向组织[47]。而在外泌体中加载其他药物,或者进行工程化修饰,能赋予它们对特定组织、受体细胞的选择性,改变体内分布,实现更高效的药物靶向递送,从而提高治疗效果。
-
有多种物理方法可以将药物加载到外泌体中,如共孵育法、电穿孔、机械挤压、超声、冻融等。其中共孵育法简单快捷,但效率低且不适合亲水性分子;电穿孔可以同时作用于亲水和疏水物质,但容易引起外泌体聚集并损害外泌体膜的完整性;机械挤压和超声载药效率高,能同时改变外泌体尺寸,但也会引起聚集和膜损伤;冻融方法简单快速,但效率低且对外泌体尺寸有很大影响。
小的亲脂性分子可以通过与外泌体共孵育被动加载到外泌体中,比如多柔比星(Dox)和紫杉醇(PTX),它们在室温下与外泌体共同孵育即可载入外泌体,其负载能力分别为PTX 7.2%,Dox 11.7%[48- 49]。除此之外,利用物理的辅助手段,可以增加生物活性大分子药物的载入。例如,Haney等[37]用外泌体装载过氧化氢酶治疗帕金森病,外泌体保护了过氧化氢酶不被降解,并使其在病变部位持续释放,从而用于治疗神经疾病。他们研究了包括机械挤压、超声、皂苷透化、冻融循环等机械处理方法对加载过氧化氢酶效率的影响,发现超声和挤压外泌体能最显著提高其过氧化氢酶的加载效率并提高其释放能力,而使用皂苷透化处理能提高其加载效率但降低其释放能力。Alvarez-Erviti等[50]将siRNA通过电穿孔方法加载到外泌体中用于治疗阿尔茨海默病,沉默β-淀粉样蛋白形成相关的BACE1基因,目标蛋白敲除效率达到62%。而Kamerkar等[51]则将沉默KrasG12D的siRNA加载到外泌体中,成功抑制了多种胰腺癌小鼠模型中的肿瘤生长,显著提高了动物的生存率。Shtam等[52-53]分别利用Lipofectamine试剂转染法和电穿孔法将siRNA导入到HeLa细胞外泌体中,而后外泌体可将siRNA转移到受体细胞内,这是一种类似于外源性纳米微粒的核酸递送方法,但转染试剂存在一定毒性,且其载入效率不如电导入方法[54]。Sterzenbach等[55]用WW标签标记目标蛋白Cre重组酶,能使其被含有L结构域的蛋白Ndfip1识别并泛素化,从而将其加载到外泌体中,因此目标蛋白的泛素化能提高外泌体的蛋白装载。
-
将药物装载于外泌体中最广泛应用的方法是以药物孵育外泌体源细胞,使得分泌的外泌体携带部分药物;或者转染源细胞,源细胞可以过表达特定的产物并将其包装到外泌体中或在外泌体膜上[13]。如利用DNA载体表达能与外泌体蛋白融合的蛋白HPV-E7和Nef,这些融合蛋白富含于外泌体中,从而提高了靶蛋白在外泌体中的特异性和负载效率[56]。通过使用miRNA表达载体或pre-miRNA,将miRNA转染到源细胞中,然后导入外泌体,过表达miR-29的HEK293T细胞所产生的外泌体抑制了胃癌的血管生成[57]。L.H.等[58]分别用PTX、依托泊苷、卡铂、伊立替康、表柔比星和米托蒽醌等孵育HepG2细胞,随后发现该细胞的外泌体与人胰腺细胞系CFPAC-1共孵育,能产生显著的抗增殖活性,外泌体通过源细胞的药物摄取实现载药。此外,刺激源细胞有时也能提高外泌体内有效物质的含量,Zhang等[59]用三种刺激物刺激THP-1单核细胞,导致其分泌的外泌体中miR-150水平升高从而增强其效能。
-
细胞来源、给药途径和剂量等许多因素都会影响外泌体在体内的生物分布。天然的外泌体通常分布到小鼠的肝脏、脾脏、肠道和肺部,与外源性纳米微粒一样,易被单核吞噬细胞系统捕获从而清除[60]。与对照小鼠相比,在巨噬细胞耗竭的小鼠中,外泌体从循环中清除的速度要慢得多[61]。为了克服这些缺点,进行工程化修饰的外泌体增加了其在被单核吞噬细胞清除之前到达靶细胞/组织的能力,从而提高药物递送效率,减少脱靶/副作用。在外泌体的天然趋向性基础上,研究人员主要通过如共价修饰、基因修饰等方法实现增加外泌体的靶向递送性能[62-63]。
-
点击化学利用炔烃和叠氮残基之间的共价相互作用形成稳定的三唑键,可用于在各种水性缓冲液(包括水、醇和二甲亚砜)中将靶向分子连接到外泌体表面[64-65]。聚乙二醇化是用支链PEG修饰外泌体表面,是使用共价连接的化学偶联方法最常见方法[66]。Kim等[67]用氨基乙基茴香酰胺-PEG(AA-PEG)修饰外泌体,AA-PEG是σ受体的靶向配体,外泌体通过AA-PEG靶向至过度表达σ受体的肺癌。Chen等[68]证明通过点击化学用神经纤毛蛋白-1靶向肽RGE修饰的外泌体可促进其在原位胶质瘤小鼠模型中对血脑屏障的渗透和胶质瘤靶向,肿瘤组织的药物积累量增加了近1.5倍,并且外泌体在肿瘤中的保留时间延长。类似地,c(RGDyK)是一种对整合素avb3具有高亲和力的肽,在缺血后脑血管内皮细胞中表达,Tian等[69]通过点击化学将其结合到MSC-Exo表面,用于治疗中风,c(RGDyK)修饰的外泌体对小鼠缺血性脑损伤区域的趋向性比对照组高11倍。而Smyth等[70]发现Azide-Fluor545荧光分子可以通过基于炔烃的交联反应附着在外泌体表面,而不会改变外泌体的大小和特性。共价键是一种非常稳定的键,但该反应不是位点特异性的,因而无法控制哪些氨基(例如,N-末端氨基、赖氨酸残基的ε-氨基)或哪些蛋白质被修饰,可能会屏蔽一些蛋白质-蛋白质相互作用从而改变外泌体的识别特性。
通过共价结合将siRNA与脂肪酸、甾醇和维生素等脂质结合物结合,可通过间接的疏水相互作用将靶分子插入外泌体膜[71]。Vandergriff等[72]将心脏干细胞衍生的外泌体通过DOPE-NHS与心脏归巢肽CHP链接,介导外泌体靶向至心脏。与DOPE一样,胆固醇因为具有疏水性也可以自组装成外泌体膜。Pi等[73]用RNA适配子或叶酸缀合的胆固醇对外泌体进行表面修饰,将siRNA和miRNA递送到相应的肿瘤部位并增强了抗肿瘤功效。
-
除了以上直接修饰外泌体的方法外,还可以通过基因修饰产生外泌体的源细胞从而间接对外泌体进行工程化修饰。与直接修饰外泌体相比,该方法在靶向物表达产量和稳定性方面具有优势[74-75]。研究人员通过转染在外泌体源细胞上表达与外泌体膜成分(如四跨膜蛋白、Lamp2b和乳黏蛋白的C1C2结构域)融合的靶向部分(如肽、受体和抗体),从而产生具有相应靶向物的外泌体[74-76]。如用狂犬病毒糖蛋白RVG修饰Lamp2B的N端,RVG与富含神经元细胞的乙酰胆碱受体特异性结合,将融合了RVG的Lamp2B转染细胞,产生了在膜外表达RVG蛋白的外泌体[50]。Kooijmans等[77]将编码抗表皮生长因子受体(EGFR)的基因转染到Neuro2A细胞内,使该细胞产生的外泌体表达EGFR,使其能与源自衰变加速因子(DAF)的糖基磷脂酰肌醇(GPI)锚定信号肽融合,从而靶向肿瘤细胞。Shimbo等[78]将合成的miR-143转入骨髓间充质干细胞,所分泌的外泌体内miR-143增加,这种过表达miR-143的外泌体递送到人骨肉瘤细胞系143B可以导致其迁移受到抑制。
-
近年来,多种外泌体-纳米粒杂合体的开发例如外泌体-脂质体、外泌体-无机纳米粒等,极大丰富了外泌体的可塑性,已被用于多种疾病的协同诊断和治疗。
-
外泌体-脂质体杂合可用于优化外泌体表面的特性,增加其载药性能和胶体稳定性、提高粒子在血液中的稳定性,通过脂质体表面修饰增加靶细胞对外泌体的摄取。Tan等[79]设计了外泌体-脂质体杂化纳米粒从而更有效地输送CRISPR-Cas9表达载体,这些杂化纳米粒被内吞后有效地抑制了MSCs中mRunx2基因和hCTNNB1基因的表达。Sato等[67, 80]等将外泌体和脂质体在液氮中反复多次冻融来产生外泌体-脂质体杂合体,与从RAW264.7细胞或HeLa细胞中分离的原始外泌体相比,外泌体-脂质体杂合体增强了与HeLa细胞的膜融合,证明了这些杂合外泌体提高了对受体细胞的靶向性。但冻融法也有缺点,它有可能改变外泌体上膜蛋白的完整性和方向性,从而削弱它们的生物功能[81]。外泌体-脂质体杂合可以提高药物的递送效率,如Piffoux等[82]使用静电作用来诱导阳离子脂质体与外泌体的融合,强阳离子电荷增强了外泌体与受体细胞的结合和细胞摄取,与游离药物或载药脂质体前体相比,膜融合的杂合外泌体将化疗药物的细胞递送效率提高了3~4倍。此外,杂合体的脂质电荷还会影响靶细胞的摄取,与阳离子脂质体杂合的外泌体相比,中性或阴离子脂质体杂合的外泌体更有可能被癌细胞系摄取[64-65]。
-
金属纳米粒子、金属氧化物纳米粒子和量子点(QD)等无机纳米粒子具有优异的物理特性,如等离子体、磁性和荧光特性等,因此利用这些纳米粒与外泌体杂合,可能会产生协同治疗效果。Khongkow等[83]用机械挤压的方法制备的外泌体和金纳米粒的杂合粒子,可以改善它们穿过血脑屏障的转运能力并特异性识别和靶向神经元细胞。Wang等[84]将装载有阿霉素的外泌体与磁性纳米粒Fe3O4杂合,使得杂合粒子可以在外部磁场诱导下靶向肿瘤部位从而产生更好的治疗效果。Cheng等[85]将肿瘤细胞外囊泡与金属-有机框架材料(MOF)杂合,利用生物膜成分保护蛋白质免受蛋白酶消化和逃避免疫系统清除,并选择性地靶向同型肿瘤部位,促进肿瘤细胞摄取和内化粒子后负载药物的自主释放。
-
外泌体是细胞释放的一类天然纳米级囊泡,可以被体内大多数细胞分泌,作为一种特殊的细胞间通讯载体,携带和传递重要的信号分子,在多种生理、病理过程中发挥着重要作用。随着以微流控芯片为代表的外泌体检测手段的不断更新,外泌体的研究为多种疾病的提前诊断和预后评估提供支持[86]。外泌体具有不均一性和异质性的特点,使不同来源的外泌体具有不同的生物学效应,不仅涉及神经系统疾病、癌症、心血管疾病、免疫反应、器官发育、组织动态平衡等,还涉及植物学和微生物学的研究内容[74]。但是外泌体的分离纯化过程依然存在一些问题,无论是离心法还是商业试剂盒都不能特异性地完全分离外泌体,从培养基中分离和纯化的外泌体仍然含有大量的非外泌体成分,如微泡和凋亡小体等功能性囊泡的存在,可能会影响外泌体医学应用的准确性和可靠性。此外,不同的细胞类型、培养条件和细胞的基因组变化也可能改变外泌体中的关键调控因子,因此需要更加准确、规范、快速、特异的分离纯化方法和液体活检技术对其质量控制标准化。作为一种内源性的纳米载体,低免疫原性和天然的靶向及归巢能力,天然的细胞间转运生物物质的机制,使外泌体作为药物传递平台具有广阔的前景。尽管多种载药方式的探索,以及工程化修饰使外泌体用于靶向药物递送被广泛研究,但如何影响外泌体的稳定性、它们的细胞进入途径和体内组织分布代谢仍有待阐明。作为高精度探针提供疾病提前诊断,具有靶向能力的药物输送体系,共同构建用于体内跟踪、预后监测和治疗的外泌体多功能平台将具有广阔的临床应用前景。
Progress on exosomes in the diagnosis and treatment of disease and drug delivery system
-
摘要: 外泌体是细胞外囊泡的一种,作为特殊的细胞间通讯介质,携带蛋白质、核酸及脂质等,在生物体内各种生理、病理过程中发挥着重要作用。作为内源性纳米囊泡,外泌体具有体循环稳定性、良好的生物相容性、对组织和细胞的特异性靶向等优点,是理想的药物递送载体。外泌体为多种疾病的诊断和预后评估提供支持,同时作为一种非常有潜力的、安全、特异性强的内源性纳米药物载体具有广阔的应用前景。本文阐述外泌体的产生机制,对其提取分离方法特点进行总结,并围绕外泌体在免疫和炎症相关疾病、心血管系统疾病、神经系统疾病、肿瘤等疾病的应用机制进行讨论,以及作为药物载体的工程化修饰和主动靶向药物递送进行综述。Abstract: As a type of extracellular vesicles, exosomes are released by living cells and contain diverse bioactive molecules, including nucleic acids, proteins, lipids and metabolites. They play an important role in various physiological and pathological processes by a special intercellular communication medium. As endogenous vesicles, exosomes also have the advantages of systemic circulation stability, good biocompatibility and specific targeting of tissues and cells, as well as they are promising candidates for drug delivery system. The production mechanism of exosomes describe was summarized, the methods of extraction and separation the application and mechanism of exosomes in immune and inflammation-related diseases, cardiovascular system diseases, nervous system diseases, tumors, etc. were reviewed. The engineering modifications of exosomes in high targeting properties based on the drug delivery were overviewed. Exosomes support the diagnosis and prognostic assessment of multiple diseases, which have broad application prospects as a very potential safe and specific endogenous nano-drug carrier.
-
Key words:
- exosome /
- targeted modification /
- targeted therapy /
- drug delivery
-
流感是一种严重的上呼吸道病毒感染,由于其高毒力和突变率,该病毒仍然是对公众健康的主要威胁。据美国疾病控制与预防中心和世界卫生组织估计,每年有多达65 万人死于季节性流感引起的呼吸道疾病[1]。传统中医药预防和治疗流感病毒导致的感染(如呼吸道肺炎和支气管炎)已成为中国临床上常规治疗策略,发挥了独特的医疗优势[2,3]。
感冒安颗粒是由金银花、连翘、板蓝根、拳参、桑叶、紫苏和荆芥等七味中药制成的复方制剂,临床应用30 多年,具有疏散风邪,解表退热功能,其预防和治疗呼吸道感染的作用已得到很好的临床验证,特别是在病毒感染初期的治疗效果尤其显著,但药效物质基础并不清楚。目前,建立了TLC法对方中的板蓝根和连翘两味药材进行特征鉴别;同时对方中各药味的共有成分绿原酸和齐墩果酸也建立了TLC鉴别方法。在含量限度方面,则建立了制剂中连翘酯苷A的HPLC法。尽管这些获批的标准已用于制剂常规质量控制,但尚需进一步进行质量评价,以期更客观、准确地反映感冒安颗粒的临床疗效。
由于中药复方复杂的化学成分,它的药理作用是通过多靶点、多途径而实现的。流感病毒感染的病理生理过程主要是病毒的直接作用和宿主免疫反应损伤(如细胞因子风暴所致的炎性损伤和ROS导致的氧化应激损伤)的结果[2,4]。为了研究感冒安颗粒临床效果的药效物质基础,我们对组方各药味的化学成分进行了文献追踪,发现方中药味含有多种黄酮类成分,如异槲皮苷、芦丁、木樨草素及木樨草苷等[5-9]。而黄酮类成分对流感病毒的作用越来越受到关注[10,11]。异槲皮苷抑制流感病毒A和B的复制,与抗病毒药amantadine或者oseltamivir合用可抑制它们导致的耐药病毒出现[12]。槲皮素与流感病毒A H1N1(A/PR/8/34)的神经酰胺酶的结合与zanamivir相当,体内研究也证实了其抗流感病毒能力,可作为抗甲型H1N1流感的有效先导化合物[13]。Zima研究认为木犀草素及其同源物是强效流感核酸内切酶抑制剂,揭示黄酮类化合物的抗流感作用[14]。鉴于此,本研究在总黄酮含量测定的基础上[15],采用HPLC-MS/MS方法建立5 种黄酮类成分的定量方法,不仅为制剂质量评价提供方法学,也为进一步研究感冒安颗粒防治流感病毒引起的呼吸道感染机制奠定物质基础。
1. 仪器与试药
1.1 仪器
Agilent Technologies
6410 Triple Quad LC/MS仪,配以Triple Quad B.02.01(B2043.12)数据处理软件(美国安捷伦公司);XS205DU电子天平(瑞士梅特勒-托利多仪器有限公司);DL-720A超声波清洗器(上海之信仪器有限公司)。1.2 试药
感冒安颗粒(本院院内制剂,批号110418、110704、111025、111121、111130、111221、120131、120201、120207、120214);芦丁对照品(中国食品药品检定研究院,供UV法测定,含量以92.5 %计,批号:100080-200707);金丝桃苷对照品(中国食品药品检定研究院,供含量测定用,含量以93.9 %计,批号:111521-201004);木犀草素对照品(中国食品药品检定研究院,供含量测定用,批号:111520-200504);槲皮素对照品(中国食品药品检定研究院,供含量测定用,批号:100081-200406);异槲皮苷对照品(成都曼斯特生物制品有限公司,纯度>99.0 %,批号:MUST-10021901);甲醇为色谱纯;甲酸为分析纯;水为蒸馏水。
2. 方法与结果
2.1 溶液制备
2.1.1 对照品溶液制备
分别取芦丁、金丝桃苷、异槲皮苷、槲皮素和木犀草素对照品各适量,用甲醇溶解,摇匀,各配制成500 μg/ml的对照品贮备液。分别精密量取5种对照品贮备液适量,稀释成浓度如下:分别含芦丁0.25、0.50、1.00、2.00、4.00、6.00、8.00 μg/ml,金丝桃苷0.002、0.008、0.020、0.032、0.044、0.056、0.080 μg/ml,异槲皮苷0.03、0.10、0.25、0.45、0.65、0.95、2.00 μg/ml,槲皮素0.02、0.10、0.20、0.30、0.40、0.60、0.70 μg/ml,木犀草素0.012、0.026、0.040、0.054、0.082、0.096、0.200 μg/ml的混合对照品溶液。
2.1.2 供试品溶液制备
按已优化的黄酮提取方法进行[15]。取样品约0.5 g,精密称定,置量瓶中,加70 %甲醇35 ml,超声提取30 min,放冷,过滤,滤液加70 %甲醇溶液定容至50 ml,摇匀,用微孔滤膜(0.45 μm)滤过,即得。
2.2 色谱-质谱条件
色谱条件:采用Kromasil C 18(4.6 mm×150 mm,5 μm,100 Å)色谱柱;甲醇(A)- 0.1 %甲酸(B)作为流动相,按0~20 min,35 % A;20~40 min,45 % A梯度洗脱。
质谱条件:电喷雾负离子化(ESI−)源:毛细管电压 3.0 kV;气体温度 350 ℃,气体流速 10 L/min,雾化气压 35 psi。多反应模式(MRM)监测。5种黄酮检测的离子对:芦丁m/z 609.1→300.1、金丝桃苷和异槲皮苷m/z 463.0→300.1、槲皮素m/z 301.0→151.0、木犀草素m/z 285.0→132.9。
2.3 方法学考察
2.3.1 检测限和定量限
采用上述色谱条件,每个待测化合物对照品用70%甲醇溶液进行系列稀释,分别以信噪比(S/N)等于3和10确定各自的检测限和定量限。结果见表1。
表 1 5种黄酮成分的线性方程、相关系数、线性范围、检测限和定量限黄酮化合物 线性方程 相关系数
r线性范围
(ng/ml)检测限
(ng/ml)定量限
(ng/ml)芦丁 Y=24 527X–162.17 0.999 7 250~8 000 0.025 0.50 金丝桃苷 Y=34 123X– 1.7381 0.999 1 2~80 0.005 0.01 异槲皮苷 Y=29 935X+1 597.80 0.999 1 30 ~2 000 0.02 0.50 槲皮素 Y=19 667X+370.71 0.999 2 20~700 0.02 0.10 木犀草素 Y=33 076X–177.98 0.999 7 12~200 0.005 0.01 2.3.2 线性范围考察
精密量取“2.1.1”项下配制的5 种黄酮成分的对照品混合溶液,照“2.2”项下进样测定,记录各待测组分的峰面积积分值。横坐标为黄酮成分质量浓度(X,ng/ml),纵坐标为峰面积(Y),进行线性回归,计算回归方程和相关系数。结果见表1,表明5 种黄酮成分在各自浓度范围内呈良好的线性关系。
2.3.3 精密度试验
精密吸取供试品溶液(批号
120131 ),照“2.2”项下操作,进样,连续测定5 次和连续测定5 d,记录各待测组分的峰面积积分值,计算日内、日间RSD。结果显示,芦丁、金丝桃苷、异槲皮苷、槲皮素和木犀草素的日内精密度RSD分别为1.09 %、1.42 %、1.69 %、0.86 %、1.27 %(n=5),日间精密度RSD分别为1.85 %、1.76 %、1.43 %、2.01 %、1.90 %(n=5),表明仪器精密度良好。2.3.4 重复性试验
取同一批号样品(批号
120131 )5 份,各0.5 g,精密称定,照“2.1.2”项下方法制备供试品溶液,照“2.2”项下操作,进样,测定峰面积积分值,并进行含量测定。结果显示,芦丁、金丝桃苷、异槲皮苷、槲皮素和木犀草素的含量分别为260.16、1.84、19.76、13.39、3.73 μg/g(n=5),RSD分别为1.51 %、1.73 %、0.90 %、1.44 %、1.68 %(n=5),表明方法的重复性良好。2.3.5 加样回收率试验
取已知含量的样品(批号120131)9 份,每份约0.5 g,精密称定,各精密加入对照品贮备液适量,使已知样品中加入的相当对照品量分别含芦丁140.00 μg、金丝桃苷0.90 μg、异槲皮苷10.00 μg、槲皮素7.00 μg、木犀草素1.90 μg的各对照品贮备液,按“2.1.2”项下的方法制备供试品溶液,照“2.2”项下操作,进样,测定峰面积积分值,计算加样回收率。结果显示,芦丁、金丝桃苷、异槲皮苷、槲皮素和木犀草素的平均加样回收率分别为:102.06%、101.60%、100.63%、102.81%、101.80%(n=9),RSD分别为1.56%、1.93%、0.67%、2.07%、1.84%(n=9)。
2.3.6 样品含量测定
取10 个批号的感冒安颗粒,分别按“2.1.2”项下的方法制备供试品溶液,照“2.2”项下操作,进样,测定峰面积积分值,计算含量,结果见表2。
表 2 不同批号感冒安颗粒含量测定结果($\bar x $ ±s, n=3)批号 芦丁 金丝桃苷 异槲皮苷 槲皮素 木犀草素 含量(μg/g) RSD(%) 含量(μg/g) RSD(%) 含量(μg/g) RSD(%) 含量(μg/g) RSD(%) 含量(μg/g) RSD(%) 110418 479.83±1.99 0.41 0.855±0.004 0.49 20.54±0.25 1.23 14.83±0.05 0.31 3.46±0.06 1.70 110704 198.98±3.01 1.52 0.596±0.006 1.05 11.79±0.20 1.68 15.31±0.13 0.86 4.51±0.01 0.19 111025 32.23±0.26 0.83 0.993±0.012 1.26 13.31±0.00 0.02 6.53±0.04 0.55 5.15±0.08 1.58 111121 69.18±1.03 1.51 0.499±0.006 1.26 13.36±0.21 1.56 8.04±0.14 1.69 5.65±0.07 1.19 111130 67.53±0.27 0.40 0.533±0.008 1.57 13.36±0.21 1.54 7.48±0.12 1.62 5.42±0.06 1.15 111221 275.38±3.61 1.31 0.291±0.002 0.73 11.44±0.02 0.21 15.74±0.02 0.14 6.51±0.02 0.24 120131 264.55±0.51 0.19 1.825±0.012 0.68 20.29±0.04 0.17 13.66±0.02 1.52 3.78±0.01 0.26 120201 239.19±1.55 0.65 0.593±0.010 1.76 18.95±0.06 0.33 18.41±0.21 1.16 3.86±0.03 0.81 120207 109.20±2.14 1.97 0.503±0.004 0.83 18.00±0.21 1.16 6.72±0.08 1.22 4.21±0.02 0.37 120214 108.93±0.59 0.54 0.461±0.002 0.45 17.67±0.10 0.01 6.32±0.08 1.19 4.73±0.01 0.11 3. 讨论
3.1 MS/MS条件的优化
为了获得良好的MS结果,优化了检测的离子模式、碎裂电压、碰撞能量等参数,以获得高灵敏度的分子离子和碎片离子。结果显示,ESI采用负离子模式可使待测的黄酮成分有更高的灵敏度。对照品混合液经产物离子扫描显示,芦丁的主要碎片为m/z:300.1、271.0;金丝桃苷和异槲皮苷的主要碎片为m/z:300.1、270.8;槲皮素的主要碎片为m/z:178.6、151.0、120.9、106.9;木犀草素的主要碎片为m/z:150.9、132.9、106.8。依据定量碎片离子选择原则,从远离母离子、裂解方式稳定、碎片离子有足够的丰度等方面进行考察,最终选择的碎裂电压、碰撞能量和定量离子如表3所示。由于金丝桃苷和异槲皮苷是结构异构体,它们有相同分子离子峰[M-H]− m/z 463,MS/MS图谱中有相同的产物离子峰m/z 300,这两个化合物通过比较两者在HPLC中的保留时间进行定位。
表 3 5种黄酮化合物的质谱检测参数黄酮化合物 母离子 产物离子 碰撞电压
(U/ V)碰撞能量
(U/ eV)芦丁 609.1 300.1 190 38 金丝桃苷 463.0 300.1 170 25 异槲皮苷 463.0 300.1 170 25 槲皮素 301.0 151.0 130 19 木犀草素 285.0 132.9 150 37 3.2 定量方法学验证
采用已优化的测定条件,感冒安颗粒中5 种黄酮类成分通过与各自标准对照品的保留时间和MS谱图的比较得以鉴别和确认,结果见图1。由于金丝桃苷、异槲皮苷和芦丁均是以槲皮素为苷元,结合不同的糖而形成,金丝桃苷与异槲皮苷还具有相同的分子量,它们的保留时间非常相近。采用HPLC-UV或二极管阵列检测的方法专属性不强,待测成分间相互干扰,很难保证测定结果的准确性。而本研究采用MS/MS-MRM模式测定,实现了准确定量的目的。
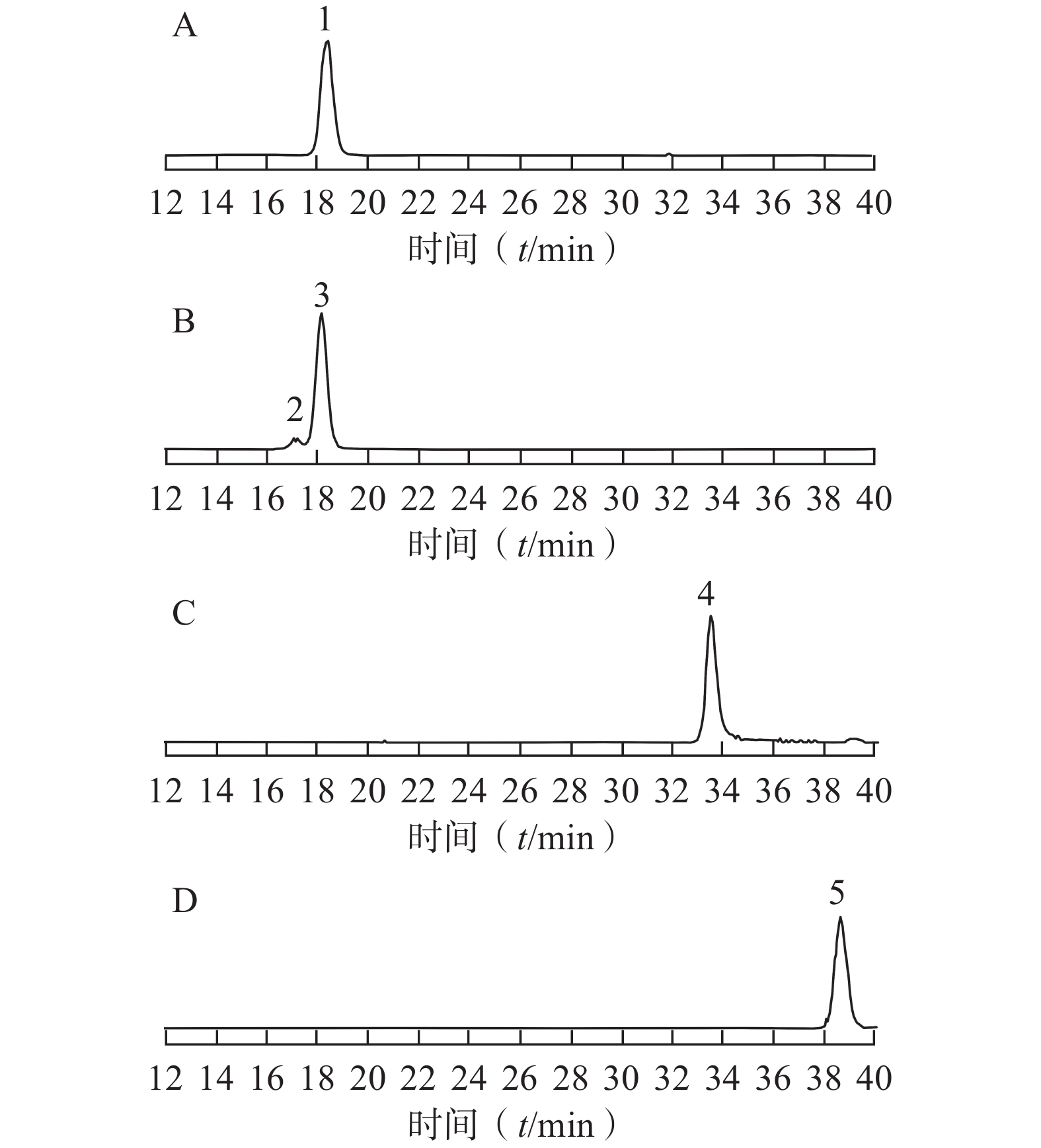 图 1 感冒安颗粒提取液5 种黄酮成分的MRM色谱图A. 芦丁的MRM色谱图;B.金丝桃苷、异槲皮素的MRM色谱图;C.槲皮素的MRM色谱图;D.木犀草素的MRM色谱图;1.芦丁;2. 金丝桃苷;3.异槲皮苷;4.槲皮素;5.木犀草素
图 1 感冒安颗粒提取液5 种黄酮成分的MRM色谱图A. 芦丁的MRM色谱图;B.金丝桃苷、异槲皮素的MRM色谱图;C.槲皮素的MRM色谱图;D.木犀草素的MRM色谱图;1.芦丁;2. 金丝桃苷;3.异槲皮苷;4.槲皮素;5.木犀草素3.3 结果分析
本研究采用经优化的超声提取法提取感冒安颗粒中黄酮成分,用所建立的LC-MS/MS方法测定了10 个批号样品,结果显示,每批样品中均为芦丁含量最高,金丝桃苷含量最低;批间样品同种黄酮成分含量存在差异。为了保证感冒安颗粒质量的稳定性,临床疗效的一致性,对其中的主要黄酮成分可以考虑设定最低限度要求。感冒安颗粒是经传统水提醇沉工艺制得的稠膏制粒而成,我们以往的研究表明,制剂中含有大量的酸性成分[16],工艺提取过程中的高温、偏低的pH值可能导致金丝桃苷、异槲皮苷、芦丁等苷类黄酮化合物发生水解反应。本研究将3种黄酮苷对照品及生产工艺获得的稠膏经2 % HCl溶液在80 ℃水浴加热处理30 min,进行HPLC-MS/MS分析,处理后的稠膏样品中金丝桃苷和芦丁含量降低,异槲皮苷和槲皮素含量增加;在3种黄酮苷对照品水解液中,均有槲皮素产生;除此之外,芦丁对照品溶液水解还出现了异槲皮苷。由此可以推测,感冒安颗粒的制备工艺可能导致黄酮类成分的水解和转化。
本研究采用HPLC-MS/MS同时测定了感冒安颗粒中5 种黄酮类成分的含量,所建立方法的专属性、灵敏度、精密度和准确性均已得到确证,达到了同时测定多种结构相似的黄酮类成分的目的,为其质量标准的建立提供了方法学依据。我们的研究已经证明感冒安颗粒中含有多种酚酸类成分[17],本研究又测定了其中的黄酮类成分。这些成分可能共同作用于流感病毒在宿主内复制的多环节,或者改善流感病毒对机体的炎性损伤,降低炎性细胞因子表达,改善氧化应激损伤,提高机体的免疫力等,充分发挥其多途径和多靶点作用优势,为其预防和治疗流感病毒所致呼吸道感染性疾病奠定物质基础。
-
表 1 外泌体的分离方法及特点
方法 原理 优势 耗时 纯度 产率 不足 超速离心法[8] 大小和密度不同的组分具有不同的沉积速度 金标准,适用于大批量样品,技术成熟 >4 h 中 低 仪器昂贵、操作繁琐耗时、产量低,可能会破坏外泌体[9] 密度梯度离心法 大小和密度不同的组分具有不同的沉积速度 高纯度,避免外泌体损伤 >16 h 高 低 前期准备、操作繁琐、耗时[10] 超滤法[11] 不同粒子粒径和相对分子质量的差异 操作简便,不需要特殊设备和试剂 <4 h 高 中 滤膜易堵塞,小粒径外泌体
易丢失[12]色谱法 不同粒子粒径和相对分子质量的差异 简单、经济,能较好保持外泌体生物功能和结构[13] <0.3 h 高 高 需要特殊的柱子和填料,存在脂蛋白污染 免疫亲和法[14] 抗体与外泌体特异性膜蛋白的相互作用 特异性分离外泌体 4~20 h 高 中 昂贵,耗时,分离效果取决于抗体的特异性 聚合物沉淀法[15] 外泌体在高亲水性聚合物影响下溶解度或分散性的变化 操作简单,适用于大体积
样品0.3~12 h 低 高 潜在污染物(提纯蛋白质聚集体或残留聚合物)  下载: 导出CSV
下载: 导出CSV
-
[1] HARDING C, HEUSER J, STAHL P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes[J]. J Cell Biol,1983,97(2):329-339. doi: 10.1083/jcb.97.2.329 [2] PAN B T, TENG K, WU C, et al. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes[J]. J Cell Biol,1985,101(3):942-948. doi: 10.1083/jcb.101.3.942 [3] GANGODA L, BOUKOURIS S, LIEM M, et al. Extracellular vesicles including exosomes are mediators of signal transduction: are they protective or pathogenic?[J]. Proteomics, 2015, 15(2-3): 260-271. [4] ISOLA A L, CHEN S. Exosomes: the messengers of health and disease[J]. Curr Neuropharmacol,2017,15(1):157-165. doi: 10.2174/1570159X14666160825160421 [5] ZHANG Y, LIU Y F, LIU H Y, et al. Exosomes: biogenesis, biologic function and clinical potential[J]. Cell Biosci,2019,9:19. doi: 10.1186/s13578-019-0282-2 [6] KAHLERT C, KALLURI R. Exosomes in tumor microenvironment influence cancer progression and metastasis[J]. J Mol Med (Berl),2013,91(4):431-437. doi: 10.1007/s00109-013-1020-6 [7] VAN NIEL G, D'ANGELO G, RAPOSO G. Shedding light on the cell biology of extracellular vesicles[J]. Nat Rev Mol Cell Biol,2018,19(4):213-228. doi: 10.1038/nrm.2017.125 [8] ZAROVNI N, CORRADO A, GUAZZI P, et al. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using immunocapture approaches[J]. Methods,2015,87:46-58. doi: 10.1016/j.ymeth.2015.05.028 [9] JEPPESEN D K, HVAM M L, PRIMDAHL-BENGTSON B, et al. Comparative analysis of discrete exosome fractions obtained by differential centrifugation[J]. J Extracell Vesicles,2014,3:25011. doi: 10.3402/jev.v3.25011 [10] GUPTA S, RAWAT S, ARORA V, et al. An improvised one-step sucrose cushion ultracentrifugation method for exosome isolation from culture supernatants of mesenchymal stem cells[J]. Stem Cell Res Ther,2018,9(1):180. doi: 10.1186/s13287-018-0923-0 [11] LI P, KASLAN M, LEE S H, et al. Progress in exosome isolation techniques[J]. Theranostics,2017,7(3):789-804. doi: 10.7150/thno.18133 [12] BUSATTO S, VILANILAM G, TICER T, et al. Tangential flow filtration for highly efficient concentration of extracellular vesicles from large volumes of fluid[J]. Cells,2018,7(12):273. doi: 10.3390/cells7120273 [13] BATRAKOVA E V, KIM M S. Using exosomes, naturally-equipped nanocarriers, for drug delivery[J]. J Control Release,2015,219:396-405. doi: 10.1016/j.jconrel.2015.07.030 [14] RUIVO C F, ADEM B, SILVA M, et al. The biology of cancer exosomes: insights and new perspectives[J]. Cancer Res,2017,77(23):6480-6488. doi: 10.1158/0008-5472.CAN-17-0994 [15] RYU K J, LEE J Y, PARK C, et al. Isolation of small extracellular vesicles from human serum using a combination of ultracentrifugation with polymer-based precipitation[J]. Ann Lab Med,2020,40(3):253-258. doi: 10.3343/alm.2020.40.3.253 [16] TÖGEL F, WEISS K, YANG Y, et al. Vasculotropic, paracrine actions of infused mesenchymal stem cells are important to the recovery from acute kidney injury[J]. Am J Physiol Renal Physiol,2007,292(5):F1626-F1635. doi: 10.1152/ajprenal.00339.2006 [17] NIU L J, ZHANG Y M, HUANG T, et al. Exosomal microRNA-155 as a biomarker for hepatic fibrosis diagnosis and progression[J]. Ann Transl Med,2021,9(2):137. doi: 10.21037/atm-20-7787 [18] CHEN L S, HUANG Y, DUAN Z X, et al. Exosomal miR-500 derived from lipopolysaccharide-treated macrophage accelerates liver fibrosis by suppressing MFN2[J]. Front Cell Dev Biol,2021,9:716209. doi: 10.3389/fcell.2021.716209 [19] RONG X, LIU J, YAO X, et al. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the Wnt/β-catenin pathway[J]. Stem Cell Res Ther,2019,10(1):98. doi: 10.1186/s13287-019-1204-2 [20] DAMANIA A, JAIMAN D, TEOTIA A K, et al. Mesenchymal stromal cell-derived exosome-rich fractionated secretome confers a hepatoprotective effect in liver injury[J]. Stem Cell Res Ther,2018,9(1):31. doi: 10.1186/s13287-017-0752-6 [21] NI Z H, KUANG L, CHEN H G, et al. The exosome-like vesicles from osteoarthritic chondrocyte enhanced mature IL-1β production of macrophages and aggravated synovitis in osteoarthritis[J]. Cell Death Dis,2019,10(7):522. doi: 10.1038/s41419-019-1739-2 [22] CASADO J G, BLÁZQUEZ R, VELA F J, et al. Mesenchymal stem cell-derived exosomes: immunomodulatory evaluation in an antigen-induced synovitis porcine model[J]. Front Vet Sci,2017,4:39. [23] QIN Y H, WANG L, GAO Z L, et al. Bone marrow stromal/stem cell-derived extracellular vesicles regulate osteoblast activity and differentiation in vitro and promote bone regeneration in vivo[J]. Sci Rep,2016,6:21961. doi: 10.1038/srep21961 [24] CHEN Y H, XUE K, ZHANG X D, et al. Exosomes derived from mature chondrocytes facilitate subcutaneous stable ectopic chondrogenesis of cartilage progenitor cells[J]. Stem Cell Res Ther,2018,9(1):318. doi: 10.1186/s13287-018-1047-2 [25] HERGENREIDER E, HEYDT S, TRÉGUER K, et al. Atheroprotective communication between endothelial cells and smooth muscle cells through miRNAs[J]. Nat Cell Biol,2012,14(3):249-256. doi: 10.1038/ncb2441 [26] ONG S G, LEE W H, HUANG M, et al. Cross talk of combined gene and cell therapy in ischemic heart disease: role of exosomal microRNA transfer[J]. Circulation, 2014, 130(11 Suppl 1): S60-S69. [27] BI S J, WANG C Y, JIN Y W, et al. Correlation between serum exosome derived miR-208a and acute coronary syndrome[J]. Int J Clin Exp Med,2015,8(3):4275-4280. [28] LAI R C, ARSLAN F, LEE M M, et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury[J]. Stem Cell Res,2010,4(3):214-222. doi: 10.1016/j.scr.2009.12.003 [29] ZOU L Y, MA X K, LIN S, et al. Bone marrow mesenchymal stem cell-derived exosomes protect against myocardial infarction by promoting autophagy[J]. Exp Ther Med,2019,18(4):2574-2582. [30] CHEN F, LI X L, ZHAO J X, et al. Bone marrow mesenchymal stem cell-derived exosomes attenuate cardiac hypertrophy and fibrosis in pressure overload induced remodeling[J]. In Vitro Cell Dev Biol Anim,2020,56(7):567-576. doi: 10.1007/s11626-020-00481-2 [31] JIANG Y, XIE H, TU W, et al. Exosomes secreted by HUVECs attenuate hypoxia/reoxygenation-induced apoptosis in neural cells by suppressing miR-21-3p[J]. Am J Transl Res,2018,10(11):3529-3541. [32] XIN H Q, LI Y, CUI Y S, et al. Systemic administration of exosomes released from mesenchymal stromal cells promote functional recovery and neurovascular plasticity after stroke in rats[J]. J Cereb Blood Flow Metab,2013,33(11):1711-1715. doi: 10.1038/jcbfm.2013.152 [33] ZHANG Y L, CHOPP M, ZHANG Z G, et al. Systemic administration of cell-free exosomes generated by human bone marrow derived mesenchymal stem cells cultured under 2D and 3D conditions improves functional recovery in rats after traumatic brain injury[J]. Neurochem Int,2017,111:69-81. doi: 10.1016/j.neuint.2016.08.003 [34] CUI G H, WU J, MOU F F, et al. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice[J]. FASEB J,2018,32(2):654-668. doi: 10.1096/fj.201700600R [35] SALA FRIGERIO C, LAU P, SALTA E, et al. Reduced expression of hsa-miR-27a-3p in CSF of patients with Alzheimer disease[J]. Neurology,2013,81(24):2103-2106. doi: 10.1212/01.wnl.0000437306.37850.22 [36] KOJIMA R, BOJAR D, RIZZI G, et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson's disease treatment[J]. Nat Commun,2018,9(1):1305. doi: 10.1038/s41467-018-03733-8 [37] HANEY M J, KLYACHKO N L, ZHAO Y L, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy[J]. J Control Release,2015,207:18-30. doi: 10.1016/j.jconrel.2015.03.033 [38] ROCCARO A M, SACCO A, MAISO P, et al. BM mesenchymal stromal cell-derived exosomes facilitate multiple myeloma progression[J]. J Clin Invest,2013,123(4):1542-1555. doi: 10.1172/JCI66517 [39] ZHOU W Y, FONG M Y, MIN Y F, et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis[J]. Cancer Cell,2014,25(4):501-515. doi: 10.1016/j.ccr.2014.03.007 [40] WOLFERS J, LOZIER A, RAPOSO G, et al. Tumor-derived exosomes are a source of shared tumor rejection antigens for CTL cross-priming[J]. Nat Med,2001,7(3):297-303. doi: 10.1038/85438 [41] ALTIERI S L, KHAN A N H, TOMASI T B. Exosomes from plasmacytoma cells as a tumor vaccine[J]. J Immunother,2004,27(4):282-288. doi: 10.1097/00002371-200407000-00004 [42] BINENBAUM Y, FRIDMAN E, YAARI Z, et al. Transfer of miRNA in macrophage-derived exosomes induces drug resistance in pancreatic adenocarcinoma[J]. Cancer Res,2018,78(18):5287-5299. doi: 10.1158/0008-5472.CAN-18-0124 [43] YANG S J, CHE S P Y, KURYWCHAK P, et al. Detection of mutant KRAS and TP53 DNA in circulating exosomes from healthy individuals and patients with pancreatic cancer[J]. Cancer Biol Ther,2017,18(3):158-165. doi: 10.1080/15384047.2017.1281499 [44] THEODORAKI M N, YERNENI S S, HOFFMANN T K, et al. Clinical significance of PD-L1+ exosomes in plasma of head and neck cancer patients[J]. Clin Cancer Res,2018,24(4):896-905. doi: 10.1158/1078-0432.CCR-17-2664 [45] LUDWIG S, FLOROS T, THEODORAKI M N, et al. Suppression of lymphocyte functions by plasma exosomes correlates with disease activity in patients with head and neck cancer[J]. Clin Cancer Res,2017,23(16):4843-4854. doi: 10.1158/1078-0432.CCR-16-2819 [46] CHEN G, HUANG A C, ZHANG W, et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response[J]. Nature,2018,560(7718):382-386. doi: 10.1038/s41586-018-0392-8 [47] PERETS N, BETZER O, SHAPIRA R, et al. Golden exosomes selectively target brain pathologies in neurodegenerative and neurodevelopmental disorders[J]. Nano Lett,2019,19(6):3422-3431. doi: 10.1021/acs.nanolett.8b04148 [48] TIAN Y H, LI S P, SONG J, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy[J]. Biomaterials,2014,35(7):2383-2390. doi: 10.1016/j.biomaterials.2013.11.083 [49] YANG T Z, MARTIN P, FOGARTY B, et al. Exosome delivered anticancer drugs across the blood-brain barrier for brain cancer therapy in Danio rerio[J]. Pharm Res,2015,32(6):2003-2014. doi: 10.1007/s11095-014-1593-y [50] ALVAREZ-ERVITI L, SEOW Y, YIN H F, et al. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes[J]. Nat Biotechnol,2011,29(4):341-345. doi: 10.1038/nbt.1807 [51] KAMERKAR S, LEBLEU V S, SUGIMOTO H, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer[J]. Nature,2017,546(7659):498-503. doi: 10.1038/nature22341 [52] SHTAM T A, KOVALEV R A, VARFOLOMEEVA E Y, et al. Exosomes are natural carriers of exogenous siRNA to human cells in vitro[J]. Cell Commun Signal,2013,11:88. doi: 10.1186/1478-811X-11-88 [53] LI S P, LIN Z X, JIANG X Y, et al. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools[J]. Acta Pharmacol Sin,2018,39(4):542-551. doi: 10.1038/aps.2017.178 [54] WANG X Y, ZHANG H Y, YANG H O, et al. Cell-derived exosomes as promising carriers for drug delivery and targeted therapy[J]. Curr Cancer Drug Targets,2018,18(4):347-354. doi: 10.2174/1568009617666170710120311 [55] STERZENBACH U, PUTZ U, LOW L H, et al. Engineered exosomes as vehicles for biologically active proteins[J]. Mol Ther,2017,25(6):1269-1278. doi: 10.1016/j.ymthe.2017.03.030 [56] MANFREDI F, DI BONITO P, ARENACCIO C, et al. Incorporation of heterologous proteins in engineered exosomes[J]. Methods Mol Biol,2016,1448:249-260. [57] ZHANG H Y, BAI M, DENG T, et al. Cell-derived microvesicles mediate the delivery of miR-29a/c to suppress angiogenesis in gastric carcinoma[J]. Cancer Lett,2016,375(2):331-339. doi: 10.1016/j.canlet.2016.03.026 [58] LV L H, WAN Y L, LIN Y, et al. Anticancer drugs cause release of exosomes with heat shock proteins from human hepatocellular carcinoma cells that elicit effective natural killer cell antitumor responses in vitro[J]. J Biol Chem,2012,287(19):15874-15885. doi: 10.1074/jbc.M112.340588 [59] ZHANG Y J, LIU D Q, CHEN X, et al. Secreted monocytic miR-150 enhances targeted endothelial cell migration[J]. Mol Cell,2010,39(1):133-144. doi: 10.1016/j.molcel.2010.06.010 [60] BERENGUER J, LAGERWEIJ T, ZHAO X W, et al. Glycosylated extracellular vesicles released by glioblastoma cells are decorated by CCL18 allowing for cellular uptake via chemokine receptor CCR8[J]. J Extracell Vesicles,2018,7(1):1446660. doi: 10.1080/20013078.2018.1446660 [61] GRANGE C, TAPPARO M, BRUNO S, et al. Biodistribution of mesenchymal stem cell-derived extracellular vesicles in a model of acute kidney injury monitored by optical imaging[J]. Int J Mol Med,2014,33(5):1055-1063. doi: 10.3892/ijmm.2014.1663 [62] RAYAMAJHI S, ARYAL S. Surface functionalization strategies of extracellular vesicles[J]. J Mater Chem B,2020,8(21):4552-4569. doi: 10.1039/D0TB00744G [63] BAEK G, CHOI H, KIM Y, et al. Mesenchymal stem cell-derived extracellular vesicles as therapeutics and as a drug delivery platform[J]. Stem Cells Transl Med,2019,8(9):880-886. doi: 10.1002/sctm.18-0226 [64] RAMASUBRAMANIAN L, KUMAR P, WANG A J. Engineering extracellular vesicles as nanotherapeutics for regenerative medicine[J]. Biomolecules,2019,10(1):48. doi: 10.3390/biom10010048 [65] VILLATA S, CANTA M, CAUDA V. EVs and bioengineering: from cellular products to engineered nanomachines[J]. Int J Mol Sci,2020,21(17):6048. doi: 10.3390/ijms21176048 [66] SUSA F, LIMONGI T, DUMONTEL B, et al. Engineered extracellular vesicles as a reliable tool in cancer nanomedicine[J]. Cancers,2019,11(12):1979. doi: 10.3390/cancers11121979 [67] KIM M S, HANEY M J, ZHAO Y L, et al. Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations[J]. Nanomedicine,2018,14(1):195-204. doi: 10.1016/j.nano.2017.09.011 [68] CHEN L K, MIAO W, TANG X Y, et al. The expression and significance of neuropilin-1 (NRP-1) on glioma cell lines and glioma tissues[J]. J Biomed Nanotechnol,2013,9(4):559-563. doi: 10.1166/jbn.2013.1624 [69] TIAN T, ZHANG H X, HE C P, et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy[J]. Biomaterials,2018,150:137-149. doi: 10.1016/j.biomaterials.2017.10.012 [70] SMYTH T, PETROVA K, PAYTON N M, et al. Surface functionalization of exosomes using click chemistry[J]. Bioconjug Chem,2014,25(10):1777-1784. doi: 10.1021/bc500291r [71] BISCANS A, HARASZTI R A, ECHEVERRIA D, et al. Hydrophobicity of lipid-conjugated siRNAs predicts productive loading to small extracellular vesicles[J]. Mol Ther,2018,26(6):1520-1528. doi: 10.1016/j.ymthe.2018.03.019 [72] VANDERGRIFF A, HUANG K, SHEN D L, et al. Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide[J]. Theranostics,2018,8(7):1869-1878. doi: 10.7150/thno.20524 [73] PI F M, BINZEL D W, LEE T J, et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression[J]. Nat Nanotechnol,2018,13(1):82-89. doi: 10.1038/s41565-017-0012-z [74] LIANG Y J, DUAN L, LU J P, et al. Engineering exosomes for targeted drug delivery[J]. Theranostics,2021,11(7):3183-3195. doi: 10.7150/thno.52570 [75] LUAN X, SANSANAPHONGPRICHA K, MYERS I, et al. Engineering exosomes as refined biological nanoplatforms for drug delivery[J]. Acta Pharmacol Sin,2017,38(6):754-763. doi: 10.1038/aps.2017.12 [76] VAKHSHITEH F, ATYABI F, OSTAD S N. Mesenchymal stem cell exosomes: a two-edged sword in cancer therapy[J]. Int J Nanomedicine,2019,14:2847-2859. doi: 10.2147/IJN.S200036 [77] KOOIJMANS S A A, ALEZA C G, ROFFLER S R, et al. Display of GPI-anchored anti-EGFR nanobodies on extracellular vesicles promotes tumour cell targeting[J]. J Extracell Vesicles,2016,5:31053. doi: 10.3402/jev.v5.31053 [78] SHIMBO K, MIYAKI S, ISHITOBI H, et al. Exosome-formed synthetic microRNA-143 is transferred to osteosarcoma cells and inhibits their migration[J]. Biochem Biophys Res Commun,2014,445(2):381-387. doi: 10.1016/j.bbrc.2014.02.007 [79] LIN Y, WU J H, GU W H, et al. Exosome-liposome hybrid nanoparticles deliver CRISPR/Cas9 system in MSCs[J]. Adv Sci (Weinh),2018,5(4):1700611. doi: 10.1002/advs.201700611 [80] SATO Y T, UMEZAKI K, SAWADA S, et al. Engineering hybrid exosomes by membrane fusion with liposomes[J]. Sci Rep,2016,6:21933. doi: 10.1038/srep21933 [81] MENTKOWSKI K I, SNITZER J D, RUSNAK S, et al. Therapeutic potential of engineered extracellular vesicles[J]. AAPS J,2018,20(3):50. doi: 10.1208/s12248-018-0211-z [82] PIFFOUX M, SILVA A K A, WILHELM C, et al. Modification of extracellular vesicles by fusion with liposomes for the design of personalized biogenic drug delivery systems[J]. ACS Nano,2018,12(7):6830-6842. doi: 10.1021/acsnano.8b02053 [83] KHONGKOW M, YATA T, BOONRUNGSIMAN S, et al. Surface modification of gold nanoparticles with neuron-targeted exosome for enhanced blood-brain barrier penetration[J]. Sci Rep,2019,9(1):8278. doi: 10.1038/s41598-019-44569-6 [84] WANG J, CHEN P, DONG Y, et al. Designer exosomes enabling tumor targeted efficient chemo/gene/photothermal therapy[J]. Biomaterials,2021,276:121056. doi: 10.1016/j.biomaterials.2021.121056 [85] CHENG G, LI W Q, HA L, et al. Self-assembly of extracellular vesicle-like metal-organic framework nanoparticles for protection and intracellular delivery of biofunctional proteins[J]. J Am Chem Soc,2018,140(23):7282-7291. doi: 10.1021/jacs.8b03584 [86] PENG H, JI W H, ZHAO R C, et al. Exosome: a significant nano-scale drug delivery carrier[J]. J Mater Chem B,2020,8(34):7591-7608. doi: 10.1039/D0TB01499K -

