

 下载:
下载:
 下载:
下载:

-
清开灵是由《温病条辨》安宫牛黄丸加减化裁而来,由珍珠母、栀子、水牛角、板蓝根、金银花五味中药以及胆酸、猪去氧胆酸、黄芩苷3种活性成分组成。具有清热解毒、镇静安神的功效,临床上常用于病毒性感冒、上呼吸道感染、急性扁桃体炎等疾病。现代研究表明,清开灵具有良好的抗病毒作用,对流感病毒H1N1、H5N1和H7N9以及2型登革热病毒等均有一定的抑制作用[1-2]。冠状病毒感染引起的肺炎是由病毒感染引起,与流感的起因及症状极其相似[3],因此,清开灵也有一定的干预冠状病毒的作用。网络药理学是近年来整合系统生物学和药理学的一门新兴学科,通过大量的生物学信息,对已有的分子生物学数据进行系统全面的分析,从蛋白、分子、基因层面揭示机体与疾病的相互作用机制,可以为多靶点药物的开发和机制研究提供依据;分子对接是通过受体的特征以及受体和药物分子之间的相互作用方式来进行药物设计的方法,通过结合能的大小判定受体与药物分子结合的能力。本文旨在通过网络药理学与分子对接技术探讨清开灵干预病毒性感冒的物质基础与作用机制,以期为今后清开灵用于临床预防和治疗病毒性感冒提供理论依据。
-
本研究采用计算系统生物学实验室的中药系统药理学数据库和分析平台 (TCMSP,http://lsp.nwu.edu.cn/tcmsp.php) 检索中药栀子、板蓝根、金银花的活性成分。因珍珠母和水牛角作为动物类药材在TCMSP数据库中未收录,笔者应用Traditional Chinese Medicines Integrated Database (TCMID,http://119.3.41.228:8000/tcmid/search/) 对珍珠母与水牛角进行活性成分检索。根据化合物口服生物利用度 (OB) 与类药性 (DL) 进行筛选,OB 阈值设为 OB≥30%,DL 阈值设为 DL≥0.18[4-5]。此外,由于黄芩苷易在体内代谢为黄芩素,胆酸易代谢为去氧胆酸,猪去氧胆酸易代谢为猪胆酸[6-8],故黄芩素、去氧胆酸、猪胆酸也被纳入活性成分范围内。采用 PubChem BioActivity Analysis Service 数据库(https://pubchem.ncbi.nlm.nih.gov/assay/assay.cgi) 来获取猪胆酸和猪去氧胆酸的靶点,其余活性成分靶点则通过TCMSP获得[9]。
-
以 “influenza”为关键词检索 GeneCard(https://www.genecards.org/)数据库,收集与病毒性感冒相关的作用靶点[3]。
-
为明确疾病靶点与药物靶点之间的关系,将两部分靶点进行交集,输入 Cytoscape软件构建化合物-靶点网络。Cytoscape 软件的核心架构是网络,每个节点是基因或活性成分,节点与节点之间的连接代表这些生物分子之间的相互作用,节点的度值代表网络中节点与节点相连的数目,度值越大说明该活性成分或靶点处于核心地位。
-
为更深层次探究靶点之间的相互作用关系,将交集所得的共同靶点上传至线上软件STRING10.5 (http://string db.org),构建蛋白相互作用网络。把物种选择为 Homosapiens,置信度设置为>0.9,其余参数保持默认设置,获取 PPI 网络。
-
利用 R Project 中的 org.Hs.eg.db 程序包(http://www.bioconductor.org/packages/release/data/annotation/html/org.Hs.eg.db.html)将基因 symbols 转变为 Entrez IDs,进而利用 clusterProfiler 程序包(3.6 版)(http://bioconductor.org/packages http://bioconductor.org/packages/release/bioc/html/clusterProfiler.html)对网络进行 GO 分析和 KEGG 通路分析。设定 P<0.05,并按照涉及的靶点数目多少进行排序,获取 GO 富集分析中分子功能、细胞组成和生物过程的前10 个条目以及 KEGG 富集的前 20 条信号通路。
-
为进一步筛选清开灵中可能干预病毒性感冒的潜在活性成分,将清开灵中的关键化合物胆酸、猪去氧胆酸、黄芩苷与“1.4”项下筛选出的度值较高的蛋白进行分子对接。从PDB数据库(https://www.rcsb.org/)下载靶蛋白RELA与JUN的三维晶体结构(PDB ID为1VJ7和2H7H,PDB格式),从Pubchem数据库(https://pubchem.ncbi.nlm.nih.gov/)下载胆酸、猪去氧胆酸与黄芩苷的三维结构(CAS号为81-25-4、83-49-8、21967-41-9,PDB格式),采用AutoDock Tools 1.5.6软件删除靶蛋白的水分子,分离配体和受体,添加非极性氢,计算Gasteiger电荷,分别保存为pdbqt格式的文件。将已经保存为PDB格式的化合物导入AutoDock Tools 1.5.6,添加原子电荷,分配原子类型,所有柔性键均默认可旋转,保存为pdbqt格式,作为对接配体。运行Auto Dock软件进行对接,采用Pymol软件可视化对接结果[10]。
-
通过TCMSP数据库检索到板蓝根活性成分169个,金银花活性成分236个,栀子活性成分98个,通过TCMID数据库检索到珍珠母活性成分9个,水牛角活性成分6个。根据OB≥30%与DL≥0.18筛选出板蓝根活性成分39个,金银花活性成分23个,栀子活性成分15个,其中MOL000098(槲皮素)、MOL000422(山奈酚)、MOL001194(亚油酸乙酯)和MOL003095(甘露醇)为栀子与金银花的共有成分,MOL000358(β-谷甾醇)和MOL000449(豆甾醇)为栀子、金银花和板蓝根的共有成分(图1中绿色节点表示活性成分,红色节点表示药物,橙色圈内节点为药物共有成分)。通过TCMSP数据库获取上述中药活性成分以及黄芩苷、黄芩素、胆酸、去氧胆酸的靶点,通过PubChem BioActivity AnalysisService 获取猪去氧胆酸以及猪胆酸的靶点,除去重复靶点,共获得靶点225个。

图 1 清开灵活性成分
-
从GeneCard 数据库中获得病毒性感冒靶点2 506个。
-
将“2.1”中的225个清开灵活性成分靶点与“2.2”中的2 506个病毒性感冒靶点输入Venny2.1软件,绘制韦恩图,两者进行交集后获得疾病-药物成分共同靶点100个,见图2。采用Cytoscape软件构建的活性成分-靶点网络,该网络共包含147个节点(47个化合物节点,100个靶点节点)和315条边,其中,红色节点表示化合物,绿色节点表示药物靶点,连接化合物与靶点的边表示它们之间有作用关系,见图3。表1列出了化合物-靶点网络中的关键节点、类别及其度值。活性成分度值排名前5位的是槲皮素、木犀草素、山奈酚、β-胡萝卜素和黄芩素,这5种化合物的节点数分别是71、31、23、18和15。靶点度值排名前5位的是前列腺素过氧化物合酶2(PTGS2)、前列腺素过氧化物合酶1(PTGS1)、丝氨酸蛋白酶1(PRSS1)、一氧化氮合酶(NOS2)和雌激素受体1(ESR1),分别能与44、31、15、14和13个化合物发生相互作用,体现了中药多成分、多靶点的特性。

图 2 清开灵化学成分作用靶点与病毒性感冒相关靶点交集分析

图 3 清开灵活性成分-靶点网络
表 1 化合物-靶点网络的关键节点及其度值
名称 类别 度值 槲皮素 活性成分 71 PTGS2 靶点 44 木犀草素 活性成分 31 PTGS1 靶点 31 山奈酚 活性成分 23 β-胡萝卜素 活性成分 18 黄芩素 活性成分 15 PRSS1 靶点 15 NOS2 靶点 14 栀子素 活性成分 14 β-谷甾醇 活性成分 13 ESR1 靶点 13 金合欢素 活性成分 12 ADRB2 靶点 10 MAPK14 靶点 8 CHRM1 靶点 8 -
应用 STRING 软件构建PPI网络(图4),在此网络图中包含88个节点,718条边,同时得到网络中关键靶点的频次 (图5)。根据“度值>均值”筛选出关键节点34个,包括:JUN、RELA、MAPK1、IL6、AKT1、MAPK14、CXCL8、APP、IL1B、MAPK8、VEGFA、FOS、EGFR、IL-4、ESR1、IL2、PRKCA、PRKCB、CASP8、CCL2、MYC、NFKBIA、CASP3、CTNNB1、IFNG、IL-10、CXCL2、RB1、ARG1、CHUK、CXCL10、MMP9、NOS2、STAT1。度值最高的是JUN,度值为29;其次为RELA,度值为27;MAPK14度值为25,IL-6度值为23。度值大的靶点提示在网络调控中起着关键作用,度值大的靶点很可能是清开灵治疗 病毒性感冒的关键靶点。

图 4 清开灵蛋白质-蛋白质相互作用网络

图 5 清开灵作用的关键靶点
-
GO富集条目(图6)生物过程主要包括:脂多糖反应、 氧化应激反应、金属离子反应、细胞脂多糖反应,分子功能包括:细胞因子受体结合、细胞因子活性、血红素结合、 磷酸酶结合,其在膜筏、膜微区、膜区、囊腔等处富集。KEGG富集信号通路(图7)主要包括:卡波济肉瘤相关疱疹病毒感染、乙型肝炎、人巨细胞病毒感染、流体剪切应力与动脉粥样硬化、糖尿病并发症的AGE-RAGE信号通路等。其中,卡波济肉瘤相关疱疹病毒感染的靶点主要包括IL-6、STAT1、p38、CASP8、Bax、CASP3、CASP9、CyrC、IKK、RB、E2F、ICAM1、Raf、ERK、AKT、IKBa、NFKB、JNK、AP-1、IL-8、COX2、VEGF和Myc;乙型肝炎的靶点主要包括:STAT1、JNK、p38、AP-1、ERK、IKK、IKB、NF-KB、PKC、Raf、STAT、IL-8、PCNA、MMP-9、Bax、Bcl2、CASP9、CASP3、Akt、SAPK、pRb和E2F(图8、图9)。

图 6 清开灵GO富集分析

图 7 清开灵KEGG富集分析(气泡图)

图 8 卡波济肉瘤相关疱疹病毒感染通路图

图 9 乙型肝炎通路图
-
一般认为配体与受体结合的稳态构象能量越低,产生的效应越强。笔者对清开灵中的胆酸、猪去氧胆酸合黄芩苷进行分子对接,对接模式见图10,其中,蓝紫色部分代表蛋白质,红黄色部分代表活性成分。胆酸、猪去氧胆酸合黄芩苷与RELA和JUN对接的结合能见表2。其中,HQG表示黄芩苷,ZQYDS表示猪去氧胆酸,DS表示胆酸。

图 10 清开灵主要成分与RELA和JUN分子对接模式
表 2 清开灵核心成分与RELA和JUN的结合能
化合物名称 化学式 相对分子质量 CAS号 结合能(kcal/mol) RELA JUN 胆酸 C24H40O5 408.57 81-25-4 –2.92 –4.12 猪去氧胆酸 C24H40O4 392.58 83-49-8 –2.69 –4.17 黄芩苷 C21H18O11 446.37 21967-41-9 –1.18 –2.62 -
清开灵由珍珠母、栀子、水牛角、板蓝根、金银花五味中药以及胆酸、猪去氧胆酸、黄芩苷3种活性成分组成。金银花中槲皮素、木犀草素、绿原酸等是抗流感病毒的主要活性成分,对甲型流感病毒、疱疹病毒及腺病毒均有一定的抑制作用[11]。栀子提取物对流感病毒感染所致小鼠的肺部炎症有明显抑制作用,在体外对流感病毒等6种病毒引起的细胞病变有明显抑制作用[12]。板蓝根乙酸乙酯部位的生物碱、有机酸和氨基酸衍生物与抗病毒活性密切相关[13]。现代网络药理学研究表明水牛角-珍珠母药对的解热作用可能与水牛角内的氨基酸代谢与珍珠母内金属离子代谢有关[14];胆酸-猪去氧胆酸药对可能通过作用于MR影响水、电解质代谢,从而发挥抗炎作用[9]。黄芩苷可以通过抑制H1N1流感病毒的中后期mRNA的转录和复制,抑制神经氨酸苷酶活性,以及抑制病毒诱导的巨噬细胞和肺上皮细胞中促炎基因的表达,从而影响 IL-6 和 IL-8 等的合成[15-16]。综上所述,现代研究表明清开灵干预病毒性感冒的机制可能是通过抗病毒、抗炎实现的。
本研究运用网络药理学研究方法共筛选出清开灵中活性成分90个,作用靶点225个,体现了中药多成分、多靶点联合作用的整体性和关联性的特点。根据“活性成分-靶点”网络结果,进一步得到排名前5的关键化合物为槲皮素、木犀草素、山奈酚、β-胡萝卜素和黄芩素。其中,槲皮素、山柰酚、木犀草素、黄芩素经研究发现也是目前抗击新冠疫情“三药三方”中连花清瘟胶囊、金花清感颗粒、清肺排毒汤等的核心化合物[17-19]。现代研究表明,槲皮素可以与F蛋白相互作用阻断病毒的黏附,发挥抗人呼吸道合胞病毒的作用[20];木犀草素可通过干扰外壳蛋白I复合物的表达,降低甲型流感病毒的含量[21];β-胡萝卜素对腺病毒转化的293细胞和人类疱疹病毒转化的 Raji细胞的生长增殖具有明显的抑制作用,对293细胞内整合的腺病毒早期基因的表达具有下调作用[22]。可见,实验研究进一步证实了清开灵的活性成分具有一定的抗病毒作用。此外,山奈酚可以减轻脂多糖导致的小鼠急性肺损伤,其机制可能是通过抑制肺组织促炎因子NF-κB、IL-1β、HO-1、PTGE、iNOS、TNF-α和COX-2mRNA的表达和促进抗炎因子HO-1mRNA的表达而发挥保护作用[23];槲皮素、木犀草素、β-胡萝卜素也具有抗炎活性[24-25]。综上可知,抗炎作用也是清开灵干预病毒性感冒的重要机制。
根据PPI网络分析结果和“活性成分-靶点”网络结果,清开灵干预病毒性感冒的主要作用靶点有前列腺素过氧化物合酶2(PTGS2)、前列腺素过氧化物合酶1(PTGS1)、丝氨酸蛋白酶1(PRSS1)、一氧化氮合酶(NOS2)、雌激素受体1(ESR1)、细胞因子(IL-6)和丝裂原活化蛋白激酶家族成员(MAPK8)等。GO功能富集分析结果发现,清开灵活性成分主要涉及氧化应激、细胞应答、炎性反应、免疫应答等生物学过程;KEGG通路富集主要涉及卡波济肉瘤相关疱疹病毒感染,乙型肝炎,人巨细胞病毒感染,流体剪切应力与动脉粥样硬化,糖尿病并发症的AGE-RAGE信号通路等通路,靶点富集数目最多的通路为病毒感染相关通路,如卡波济肉瘤相关疱疹病毒感染、乙型肝炎、人巨细胞病毒感染等,推测清开灵对病毒引起的感染具有一定的干预作用。现代研究表明,金银花中的绿原酸可以抑制流感病毒结构蛋白神经氨酸酶,从而阻断子代病毒的释放与传播;板蓝根中的橙黄胡椒酰胺酯可降低RNA聚合酶活性,同时可抑制NF-κB因子的转录水平;黄芩中的黄芩苷不仅可以抑制神经氨酸酶还可抑制流感病毒RNA聚合酶[26]。此外,以上通路均涉及炎症基因,如COX-2、IL-1β、IL-6、IFN-γ等,同时,TNF信号通路与IL-17信号通路在KEGG富集分析中亦有较高的相关度:TNF信号通路在抗病毒、介导能量代谢、炎症反应、血管生成以及维持干细胞特性等方面发挥重要作用,IL-17信号通路与免疫反应及炎症反应的发生与发展密切相关[10]。综上所述,清开灵可能通过抗病毒、抗炎途径起到干预病毒性感冒的作用。
胆酸、猪去氧胆酸以及黄芩苷作为清开灵中的主要成分,JUN与RELA为PPI网络中的关键蛋白。分子对接结果显示,这3种活性成分与JUN和RELA均有一定的结合能力。受体内代谢因素影响,如黄芩苷会代谢为黄芩素,胆酸、猪去氧胆酸亦会在体内代谢为去氧胆酸和猪胆酸。以黄芩苷为例,从数据库的靶点数目来看,黄芩苷的靶点数目不如黄芩素,笔者推测黄芩素多靶点的原因可能是其结构上更易与靶点蛋白结合。而清开灵中应用黄芩苷而不用黄芩素可能是因为黄芩苷不易产生消化道副作用、成本低或者与药动学有关,但实际情况仍需后续探究。
急性病毒性感冒属于中医瘟疫的范畴,具有强烈的传染性与流行性,如SARS与COVID-19均属于瘟疫,治疗上大多应用清热解毒药。清开灵中金银花、栀子、板蓝根、水牛角、珍珠母均为寒凉之品,黄芩与猪胆汁在中医学中亦属寒凉,故笔者推测黄芩中提取出的黄芩苷与猪胆汁中提取出的胆酸和猪去氧胆酸亦属寒凉,根据中医理论推测其干预病毒性感冒具有一定作用。临床研究数据显示,清开灵联合奥司他韦可以提高小儿咽部甲型流感病毒的清除率[27],与儿童清肺口服液联用可治疗小儿呼吸道合胞病毒性肺炎[28],联合利巴韦林可治疗流行性感冒[29]。综上分析,清开灵用于病毒性感冒的治疗可能与其抗病毒、抑制炎症反应相关,进一步证实了网络药理学分析结果的合理性。但近些年对清开灵注射液的安全性讨论层出不穷,使得人们对该药物望而却步[30];而且中医学认为该药苦寒败胃,不能够长期应用,故此,在临床上我们仍需辨证论治。
Mechanism of Qingkailing on influenza based on network pharmacology and molecular docking
-
摘要:
目的 探索清开灵干预病毒性感冒的潜在作用机制,为其临床用药提供理论指导。 方法 通过TCMSP、TCMID和Pubchem数据库获得清开灵的活性成分及作用靶点;通过GeneCards数据库获取病毒性感冒的相关靶点,并采用交集法筛选出与清开灵活性成分作用的共同靶点,运用Cytoscape构建“活性成分-靶点”网络,并通过String数据库构建PPI网络;通过Bioconductor数据库进行KEGG通路和GO功能富集分析,运用R软件将结果进行可视化;采用Auto Dock Tools进行分子对接研究。 结果 从清开灵中共筛选出潜在活性成分90个,对应靶点225个;PPI网络分析获得清开灵干预病毒性感冒的关键作用靶点34个;GO及KEGG富集分析得出:清开灵干预病毒性感冒的作用机制主要与抗炎、抗病毒有关;分子对接结果显示:清开灵中胆酸、猪去氧胆酸、黄芩苷与RELA和JUN具有一定的亲合力。 结论 清开灵含有的活性成分可作用于JUN、RELA、MAPK1、IL-6、AKT1等靶点,调节多条信号通路,从而发挥对病毒性感冒的干预作用。 Abstract:Objective To explore the potential mechanism of Qingkailing (QKL) on influenza, and to provide a theoretical basis for the clinical application of QKL. Methods TCMSP, TCMID, and PubChem databases were used to search for the active ingredients and action targets of QKL. GeneCards database was used to search for the targets of influenza. The intersection method was used to obtain the targets related to the therapeutic effects of QKL. Cytoscape software was applied for the construction of active compounds-targets network map. Protein-protein interaction network was constructed by STRING database. Gene ontology functional enrichment analysis and KEGG pathway enrichment analysis were conducted by Bioconductor database and R software. Auto Dock Tools were used for molecular docking. Results Total 90 potential active components were identified from QKL with the corresponding 225 targets. PPI network analysis showed that there were 34 key targets intervening influenza by QKL. GO and KEGG showed that the mechanism of QKL intervention on influenza was related to anti-inflammatory and antiviral. The results of molecular docking showed that cholic acid, hyodeoxycholic acid and baicalin had affinity with RELA and JUN. Conclusion The active ingredients of QKL target on JUN, RELA, MAPK1, IL6 and AKT1 to regulate multiple signal pathways, and play an intervention role in influenza. -
Key words:
- Qingkailing /
- network pharmacology /
- molecular docking /
- antivirus
-
啤酒花(Humulus lupulus L.)别名忽布、香蛇麻、蛇麻花、啤瓦古丽(维吾尔语),为桑科葎草属植物,是我国新疆药食兼用的特色资源植物。同时,啤酒花还是啤酒酿造的重要原料之一,其不仅赋予了啤酒独特的风味,还延长了啤酒的保质期。人类使用啤酒花已有2000多年的历史,早在公元前2世纪,古巴比伦就曾栽培使用啤酒花。13世纪,啤酒花开始作为草药使用。1516年,德国颁布法令,将啤酒花限定为啤酒的唯一苦味添加剂[1]。啤酒花的花序中含有黄酮类、树脂类、多酚、多糖等多种化学成分,这些成分使其具有抗菌、抗肿瘤、抗氧化、降血糖、降血压,以及雌激素样等药理作用[2]。近年来,啤酒花在抗骨质疏松领域研究较多,展现出广阔的前景,现就有关情况作概要介绍。
1. 化学成分
1.1 树脂类
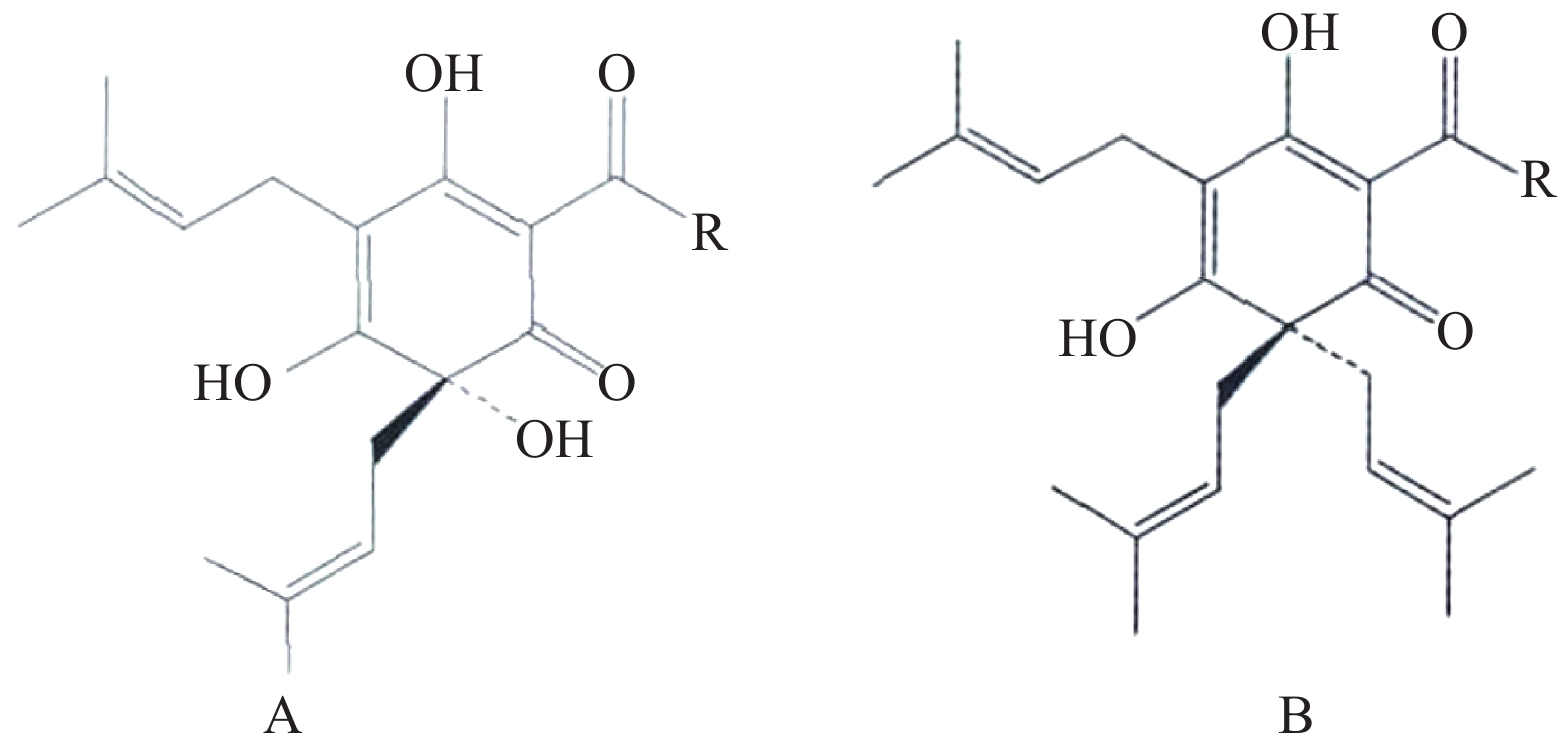
树脂类化合物为啤酒花中的主要成分,具有广泛的生物活性。欧洲酿造协会(EBC)根据不同有机溶剂中树脂类成分的溶解度差异,将其分为软树脂和硬树脂[3]。其中,α-酸和β-酸是啤酒花中最具代表性的软树脂类成分,也是啤酒花独特味道的主要来源。α-酸主要包括葎草酮(图1A)及其同系物,β-酸主要包括蛇麻酮(图1B)及其同系物[4]。在啤酒花中,β-酸含量较α-酸低,易被氧化形成β-软树脂。α-酸在一定条件下易转化为异α-酸,此类成分为啤酒中的主要苦味成分[5]。
1.2 黄酮类
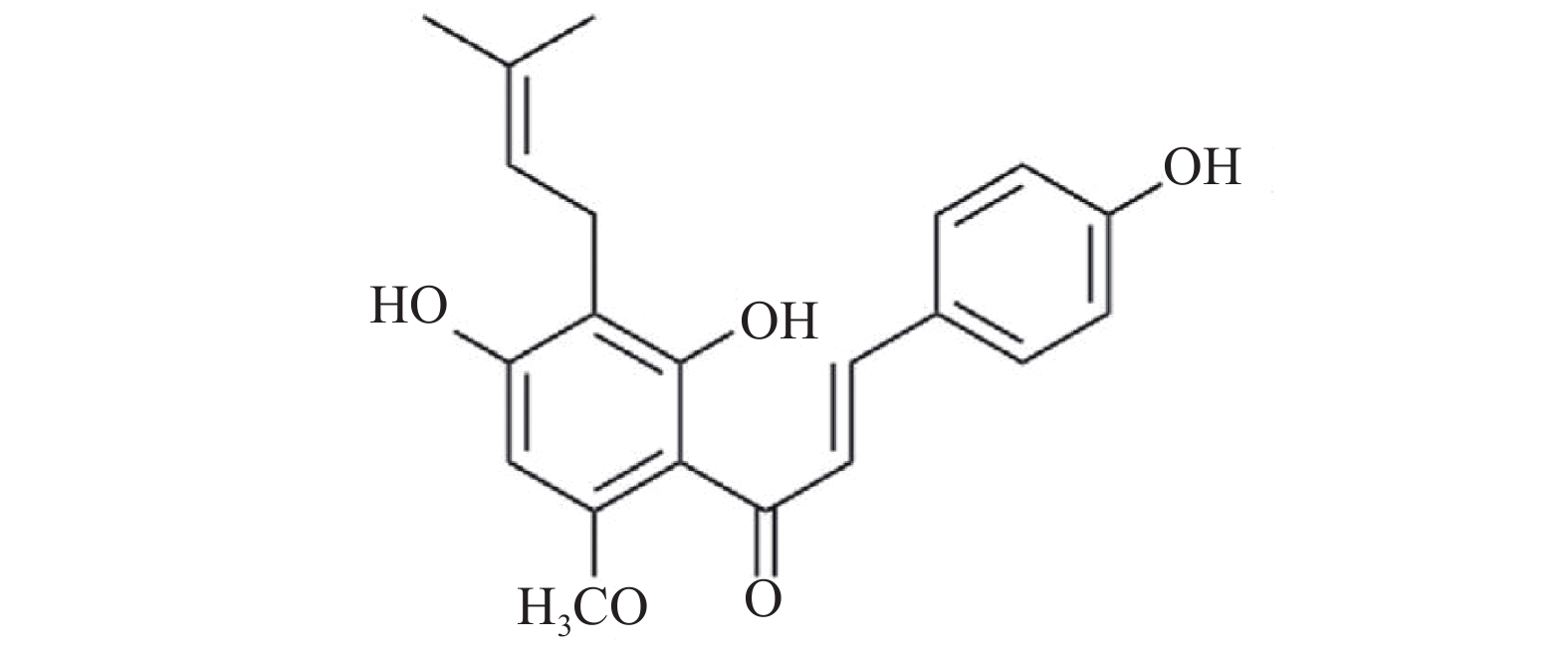
以黄腐酚(图2)为代表的黄酮类成分是啤酒花中重要的化学成分。根据母核结构的不同,可将其分为黄酮类、查尔酮类及黄烷类[6]。黄腐酚为啤酒花特有的异戊烯基类黄酮,最早被Power等[7]分离鉴定得到,主要集中在啤酒花蛇麻腺中,目前因其广泛的药理活性而备受关注。
1.3 挥发油类
啤酒花中的挥发油类成分均由啤酒花的蛇麻腺分泌,是啤酒花香味的来源。啤酒花挥发油主要含有石竹烯、香叶烯、葎草烯、法呢烯等及其脂、酮、醇类化合物[8]。早期研究普遍认为,萜烯类在啤酒花风味中起关键作用,但近年来研究表明,亲水性较强的萜烯醇类对啤酒花风味贡献更为突出[9]。
2. 抗骨质疏松作用
骨质疏松症是以骨量降低和骨微结构破坏为特点的全身性骨代谢疾病。在欧洲,啤酒花提取物用于治疗绝经后骨质疏松症。近年来,啤酒花在防治骨质疏松方面的作用受到广泛关注,其可能通过发挥雌激素样作用、缓解氧化损伤、调节骨形成-骨吸收平衡等途径维持骨稳态,来防治骨质疏松。
2.1 雌激素样作用
雌激素缺失是骨质疏松发病的主要因素之一[10]。啤酒花中的黄酮类成分8-异戊二烯基柚皮素(8-PN),是脱甲基黄腐酚的一种异构物,也是目前分离得到的最有效的植物雌激素[11]。早在2002年,Miligan等[12]即发现天然和人工合成的8-PN在人雌激素受体转染的酵母菌以及雌激素反应的人Ishikawa Var-I细胞中均显示出相似的生物活性,8-PN与两种构型的雌激素受体(ER-α、ER-β)均展现出良好的结合能力。体外筛选实验发现,其雌激素活性高于包括香豆素在内的多种常用植物雌激素。作为啤酒花中最具代表性的黄酮类成分黄腐酚同样具有显著的植物雌激素样作用。研究发现,在去卵巢小鼠中,30和90 mg/(kg·d)的黄腐酚均可显著抑制去卵巢小鼠雌激素缺失所致的体重增加,提高雌激素(E2)水平,抑制碱性磷酸酶(ALP)、抗酒石酸酸性磷酸酶(TRAP)等骨转换指标的高表达,并改善去卵巢小鼠的骨微结构破坏,增强骨密度,防治骨质空洞[13]。
啤酒花中的多种树脂类成分同样具有雌激素样作用。Holick等[14]在为期14周的临床试验中发现,树脂类成分、小檗碱、维生素D和维生素K配伍应用,可调节绝经后妇女的骨代谢水平,降低骨转换标志物骨钙素(OCN),并显著提高患者血清25-羟基维生素D含量,减少骨质丢失。Keiler等[15]以去卵巢大鼠为研究对象,观察标准酒花提取物对大鼠骨丢失的防治作用,结果显示,标准酒花提取物可显著减少大鼠胫骨干骺端破骨细胞的数量,并防止雌二醇消耗导致的骨小梁厚度减少,防治雌激素缺失所致的骨质疏松。
2.2 缓解氧化损伤
雌激素或雄激素缺失会降低骨骼防御氧化应激的能力,从而造成骨质流失,因此,高氧化应激水平与性激素缺乏同为诱发骨质疏松的重要原因[16]。黄酮类化合物大多有酚羟基以及较大的空间位阻,因此,多具有不同程度的抗氧化活性。吴婕等[17] 在研究黄腐酚与其他食品中成分的抗氧化作用时发现,黄腐酚与食品酸味剂柠檬酸、柠檬酸钠、维生素C对DPPH体系均存在协同抗氧化活性。Suh等[18]发现黄腐酚可通过激活氧化应激的关键通路Nrf2来降低MC3T3-E1成骨细胞的氧化损伤,同样证实了黄腐酚具有抗氧化作用。
2.3 维持骨稳态
成骨细胞的骨形成与破骨细胞的骨吸收在骨代谢中共同发挥作用,以维持骨稳态[19]。研究表明,在成骨细胞MC3T3-E1水平上,黄腐酚可显著上调ALP和成骨标记基因骨形成蛋白(BMP-2)、骨涎蛋白(BSP)的表达,并通过调控p38 MAPK和ERK信号通路相关机制,激活转录因子RUNX2,促进骨形成[20]。本课题组前期研究发现,黄腐酚既可以促进原代成骨细胞的增殖、ALP活性以及骨矿化水平,又可以提高骨形成相关蛋白BSP、BMP-2 和骨桥蛋白(OPN)的表达水平[13]。在抑制破骨细胞骨吸收方面,黄腐酚可通过破坏RANK与TRAF6之间的结合,抑制破骨细胞生成过程中NF-κB和Ca2+/NFATc1信号通路,并抑制破骨细胞生成相关标记基因,如组织蛋白酶K(CtsK)、活化T细胞核因子1(NFATc1)以及TRAP的表达,从而抑制骨吸收[21]。
啤酒花树脂类成分同样可以调节骨代谢平衡。蛇麻酮可显著促进成骨细胞增殖,提高ALP活性,促进骨矿化结节,并促进骨形成相关蛋白OCN、BSP和BMP-2的表达。葎草酮也可显著促进成骨细胞活性,并提高骨形成相关蛋白OCN、OPN、BSP和BMP-2的表达。在破骨细胞水平上,蛇麻酮和葎草酮均可降低破骨细胞数目,抑制破骨相关蛋白CtsK、金属基质蛋白酶9(MMP-9)的表达[22]。此外,本课题组前期还发现啤酒花乙醇提取物可显著促进成骨细胞的增殖、ALP活性及骨矿化结节,促进骨形成相关蛋白OPN和BMP-2的表达;并显著抑制破骨相关蛋白TRAP、CtsK、和MMP-9的表达,以维持骨代谢平衡,防止骨吸收大于骨形成所致的骨质流失[23]。
3. 相关产品及应用
啤酒酿制是啤酒花最为传统的一种应用方式,人体中的黄腐酚及相关异戊二烯黄酮类成分主要通过喝啤酒摄入[24]。近年来,啤酒花作为一种特色中药,其药食两用的特性得到了越来越多的关注。相关的健康产品,如美国西楚(Citra)、捷克萨兹(Saaz)的啤酒花颗粒、澳大利亚澳萃维(Nature's Way)啤酒花胶囊、芬兰麦诺美(Menomax)啤酒花浓缩片等层出不穷。现代研究已发现一些啤酒花相关产品在抗骨质疏松方面具有良好的活性。Ban等[25]采用去卵巢大鼠骨质疏松模型,研究Lifenol®的啤酒花提取物防治骨质疏松的作用,结果发现,该产品可显著改善去卵巢所致的大鼠体重增加,调节血脂和脂肪聚集,降低血流速度,改善大鼠潮热,并显著增加大鼠股骨的骨密度,改善骨质疏松。有直接证据表明,喝啤酒可以预防骨质疏松症。Kondo[26]以去卵巢大鼠为骨质疏松模型,研究啤酒对骨质疏松症的影响。结果发现,啤酒能显著抑制去卵巢引起的大鼠股骨骨丢失,且这种抑制作用在单用酒精或不加啤酒花酿造的啤酒中均没有呈现,表明啤酒中抗骨质疏松的活性成分来自啤酒花。此外,Ferk等[27]发现,人类连续饮用黄腐酚饮料14 d后,体内的氧化性嘌呤含量显著降低,氧化损伤程度得到缓解,且血清雌激素及骨钙素水平相对下降,骨代谢紊乱得到显著改善。
4. 我国啤酒花资源现状
学术界对啤酒花的起源地尚无定论,部分学者认为,啤酒花起源于中国[28-29]。在我国,啤酒花的发源地位于黑龙江省东南部的尚志市。1960年,国家轻工业部决定把新疆农场建成国家啤酒花生产基地,并从山东青岛、东北一面坡引进啤酒花。后经过40年的努力,啤酒花种植面积已达185.2公顷,成为新疆地区的主要经济作物之一。
我国的野生啤酒花种群主要分布在新疆的天山和阿尔泰山山脉附近。新疆地区由于光照充足,昼夜温差大,非常适宜啤酒花种植业的发展,并逐渐形成了范围广、规模大、生境类型多,且其成熟期差异明显的野生种质[3]。然而,近些年,啤酒花的资源开发及生产状况不容乐观。由于新疆地区种植的啤酒花主要是从美国、德国引进的品种,在异地栽培时间过长,生长性能下降,病虫害严重,品种退化严重,导致药材品质下降。再就目前常见的优质香型、香型、苦型和高α-酸型啤酒花的不同品种来看,α-酸含量高者大于8%(高α-酸型),低者3%~4%(苦型、香型),α-酸与β-酸比值高者大于2.0(高α-酸型),低者低于1.0(优质香型)。由此看来,重点活性成分苦味酸类在不同类型啤酒花中的含量、比例组成差异极大[4]。这就使得我们应加强从源头上的育种研究,改进抗病性,增加产量和苦味酸类成分的含量,利用栽培品种和育种系(或野生啤酒花)在合适条件下复合优良性状。
5. 展望
现代药理学研究分别从体内和体外实验明确了啤酒花及其活性成分的抗骨质疏松作用。黄腐酚、蛇麻酮和葎草酮均能够促进骨形成和抑制骨吸收,但其抗骨质疏松的深层次机制仍有待阐明。啤酒花对女性绝经后骨质疏松具有防治作用,但对老年性骨质疏松的作用尚不明确。因此,笔者认为阐明啤酒花及其活性成分的作用机制,既是为啤酒花临床应用及转化提供理论基础,亦可为拓展啤酒花的应用范围提供依据。此外,我国啤酒花资源丰富,但种质资源、遗传背景、亲缘关系不清,导致药材品质参差不齐。因此,摸清啤酒花种质资源家底,构建种质资源库;探讨不同基因在种群中的分布频率,弄清种群间的亲缘关系,建立药材的质量标准,方能为啤酒花活性研究和开发提供品质保障。
-
表 1 化合物-靶点网络的关键节点及其度值
名称 类别 度值 槲皮素 活性成分 71 PTGS2 靶点 44 木犀草素 活性成分 31 PTGS1 靶点 31 山奈酚 活性成分 23 β-胡萝卜素 活性成分 18 黄芩素 活性成分 15 PRSS1 靶点 15 NOS2 靶点 14 栀子素 活性成分 14 β-谷甾醇 活性成分 13 ESR1 靶点 13 金合欢素 活性成分 12 ADRB2 靶点 10 MAPK14 靶点 8 CHRM1 靶点 8  下载: 导出CSV
下载: 导出CSV
表 2 清开灵核心成分与RELA和JUN的结合能
化合物名称 化学式 相对分子质量 CAS号 结合能(kcal/mol) RELA JUN 胆酸 C24H40O5 408.57 81-25-4 –2.92 –4.12 猪去氧胆酸 C24H40O4 392.58 83-49-8 –2.69 –4.17 黄芩苷 C21H18O11 446.37 21967-41-9 –1.18 –2.62
下载: 导出CSV
-
[1] 赵利华, 陈全姣. 清开灵注射液体对H1N1、H5N1和H7N9流感病毒的作用研究[J]. 中药新药与临床药理, 2015, 26(5):644-648. [2] 范东瀛, 高娜, 安静, 等. 清开灵注射液体外抗2型登革病毒的作用[J]. 微生物学免疫学进展, 2017, 45(5):34-40. [3] 俞婷, 刘新娟, 刘力, 等. 基于网络药理学的荆银颗粒治疗病毒性感冒作用机制研究[J]. 中成药, 2020, 42(2):456-461. doi: 10.3969/j.issn.1001-1528.2020.02.034 [4] TAO W Y, XU X, WANG X, et al. Network pharmacology-based prediction of the active ingredients and potential targets of Chinese herbal Radix Curcumae formula for application to cardiovascular disease[J]. J Ethnopharmacol,2013,145(1):1-10. doi: 10.1016/j.jep.2012.09.051 [5] XU X, ZHANG W X, HUANG C, et al. A novel chemometric method for the prediction of human oral bioavailability[J]. Int J Mol Sci,2012,13(6)-6982. [6] 解立科, 田小亭, 郭小珍, 等. 黄芩素与黄芩苷微生物和肝脏代谢异同研究[J/OL]. 中成药: 1-8(2020-04-07)[2020-05-03]. http://kns.cnki.net/kcms/detail/31.1368.R.20200407.1545.002.html. [7] WAHLSTRÖM A, SAYIN, MARSCHALL H U, et al. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism[J]. Cell Metab,2016,24(1):41-50. doi: 10.1016/j.cmet.2016.05.005 [8] DE AGUIAR VALLIM T Q, TARLING E J, EDWARDS P A. Pleiotropic roles of bile acids in metabolism[J]. Cell Metab,2013,17(5):657-669. doi: 10.1016/j.cmet.2013.03.013 [9] 吴嘉瑞, 蔺梦娟, 刘鑫馗. 基于网络药理学的“胆酸-猪去氧胆酸”作用机制研究[J]. 中国医院用药评价与分析, 2018, 18(1):7-11. [10] 郭盛, 武文星, 谢红, 等. 基于网络药理学与分子对接技术的补肺活血胶囊用于新型冠状病毒肺炎(COVID-19)恢复期治疗的分子机制研究[J/OL]. 中草药: 1-10(2020-04-27)[2020-05-04]. http://kns.cnki.net/kcms/detail/12.1108.r.20200427.1221.008.html. [11] 路俊仙, 梁瑞雪, 林慧彬. 金银花抗流感病毒作用研究进展[J]. 现代中药研究与实践, 2018, 32(5):77-81. [12] 王意忠, 崔晓兰, 高英杰, 等. 栀子提取物抗病毒试验研究[J]. 中国中药杂志, 2006, 31(14):1176-1178. doi: 10.3321/j.issn:1001-5302.2006.14.013 [13] 许会芹, 何立巍, 侯宪邦. 板蓝根乙酸乙酯部位抗病毒活性组分及相关化学成分研究[J]. 南京中医药大学学报, 2019, 35(4):465-470. [14] 吴嘉瑞, 蔺梦娟, 刘鑫馗. 基于网络药理学的“水牛角-珍珠母”药对作用机制研究[J]. 中国医院用药评价与分析, 2018, 18(1):18-22. [15] SU Z Z, DOU J, XU Z P, et al. A novel inhibitory mechanism of baicalein on influenza A/FM1/1/47 (H1N1) virus: interference with mid-late mRNA synthesis in cell culture[J]. Chin J Nat Med,2012,10(6):415-420. [16] HOUR M J, HUANG S H, CHANG C Y, et al. Baicalein, ethyl acetate, and chloroform extracts of Scutellaria baicalensis inhibit the neuraminidase activity of pandemic 2009 H1N1 and seasonal influenza A viruses[J]. Evid Based Complement Alternat Med,2013,2013:750803. [17] 吉米丽汗·司马依, 买买提明·努尔买买提, 艾尼瓦尔·吾买尔, 等. 基于网络药理学及分子对接探索金花清感颗粒辅助治疗新型冠状病毒肺炎(COVID-19)活性成分研究[J/OL]. 中药材: 1-10(2020-03-24)[2020-05-05]. http://kns.cnki.net/kcms/detail/44.1286.R.20200323.1926.002.html. [18] 王法财, 沈炳香, 何春远, 等. 连花清瘟颗粒对新型冠状病毒肺炎的临床疗效及其机制的网络药理学研究[J/OL]. 中药药理与临床: 1-22(2020-03-18)[2020-05-05]. https://doi.org/10.13412/j.cnki.zyyl.20200318.001. [19] 徐天馥, 贺成功, 杨坤. 基于网络药理学清肺排毒汤治疗新冠肺炎的物质基础及作用机制研究[J/OL]. 天然产物研究与开发: 1-12(2020-04-14)[2020-05-05]. http://kns.cnki.net/kcms/detail/51.1335.Q.20200413.1918.018.html. [20] LOPES B R P, DA COSTA M F, GENOVA RIBEIRO A, et al. Quercetin pentaacetate inhibits in vitro human respiratory syncytial virus adhesion[J]. Virus Res,2020,276:197805. doi: 10.1016/j.virusres.2019.197805 [21] YAN H Y, MA L L, WANG H Q, et al. Luteolin decreases the yield of influenza A virus in vitro by interfering with the coat protein I complex expression[J]. J Nat Med,2019,73(3):487-496. doi: 10.1007/s11418-019-01287-7 [22] 刘辉. β-胡萝卜素对病毒转化细胞的影响[J]. 第四军医大学学报, 2002(6):514-516. doi: 10.3321/j.issn:1000-2790.2002.06.009 [23] 赵妍. 山奈酚对脂多糖诱导小鼠急性肺损伤的保护作用[D]. 哈尔滨: 东北农业大学, 2013. [24] CHO S O, KIM M H, KIM H. Β-carotene inhibits activation of NF-κB, activator protein-1, and STAT3 and regulates abnormal expression of some adipokines in 3T3-L1 adipocytes[J]. J Cancer Prev,2018,23(1):37-43. doi: 10.15430/JCP.2018.23.1.37 [25] 李惠香, 张倩, 柳亚男, 等. 木犀草素与木犀草苷的抗炎活性对比研究[J]. 烟台大学学报(自然科学与工程版), 2018, 31(2):114-120. [26] 周漪波, 窦文渊, 段星春, 等. 靶向流感病毒复制的中药研究进展[J]. 国际药学研究杂志, 2019, 46(5):332-338. [27] 林萌, 熊丽君, 陈贤娥, 等. 国产奥司他韦联合清开灵颗粒治疗小儿甲型流感的疗效分析[J]. 福建医药杂志, 2019, 41(1):74-76. doi: 10.3969/j.issn.1002-2600.2019.01.029 [28] 杜林波. 清开灵注射液与儿童清肺口服液联用治疗小儿呼吸道合胞病毒性肺炎痰热闭肺证的临床评价[J]. 中国社区医师, 2018, 34(35):98-99. doi: 10.3969/j.issn.1007-614x.2018.35.056 [29] 于力, 冯帅儒. 利巴韦林注射液联合清开灵注射液治疗流行性感冒临床疗效研究[J]. 世界最新医学信息文摘(电子版), 2017, 17(7):78-78, 81. [30] 张真真, 梁艳, 崔瑛, 等. 清开灵注射液的临床应用及安全性研究进展[J]. 吉林中医药, 2017, 37(3):320-324. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 9297
- HTML全文浏览量: 3635
- PDF下载量: 121
- 被引次数: 0
