

 下载:
下载:

 下载:
下载:

-
胰腺癌是一种病死率极高的消化道恶性肿瘤 [1]。超过80%以上的患者一旦确诊即是晚期,手术难以根治,需在术后进行辅助化疗、放疗、对症支持治疗等 [2]。盐酸吉西他滨(hcGEM)是治疗胰腺癌的一线化疗药。由于存在半衰期短、产生耐药性及不可避免的毒副作用等问题,其疗效不尽如人意[3]。因此,hcGEM的临床应用需要联合化疗来提高疗效[4]。
铁死亡是一种铁依赖的非凋亡性细胞死亡形式,针对铁死亡的治疗策略可能为传统疗法难以攻克的癌症提供新的治疗思路 [5]。埃拉斯汀 (Era)作为一种高效持久的铁死亡诱导剂,它可以激活多种信号通路来触发癌细胞的铁死亡 [6]。柳氮磺胺吡啶(SASP)是一种能抑制铁死亡相关的胱氨酸-谷氨酸逆向转运蛋白的抗炎药,可通过降低癌细胞对胱氨酸的摄取以及胞内谷胱甘肽水平来抑制胰腺癌细胞的生长[7]。青蒿琥酯(Art)是一种青蒿素的衍生物,除用作抗疟治疗外,可通过促进铁蛋白吞噬来增加细胞内游离铁水平,进而引发癌细胞的铁死亡[8]。铁死亡诱导剂与盐酸吉西他滨联合应用可能是胰腺癌治疗的潜在策略[9]。
本研究分别考察Era、SASP和Art这三种铁死亡诱导剂单独或联合hcGEM使用,对人胰腺癌PANC-1细胞的增殖抑制作用,以期发现具有潜在协同抑制的联合方案,为今后开发胰腺癌新疗法奠定基础。
-
盐酸吉西他滨、柳氮磺胺吡啶、埃拉斯汀(美国MCE公司);青蒿琥酯(上海泰坦科技股份有限公司);胎牛血清、青链霉素、胰酶(以色列BI公司);DMEM高糖培养基、PBS缓冲液(上海泰坦科技股份有限公司),CCK- 8细胞毒性试剂盒(日本同仁化学研究所)。
-
人胰腺癌PANC-1细胞,来源于海军军医大学第一附属医院消化内科,冻存复苏后培养于含10%胎牛血清的DMEM高糖培养基。
-
人胰腺癌PANC-1细胞用DMEM完全培养基(含10 %胎牛血清,1 %青-链霉素)于5 % CO2、37 ℃的恒温培养箱中培养。待细胞融合度达80 %~90 %时,移除旧培养基,PBS缓冲液清洗2遍后加入适当体积的胰蛋白酶消化3 min。待大部分细胞镜下变圆,小部分细胞脱落时,加入2倍胰酶体积的完全培养基终止消化,充分吹打使细胞脱离。将细胞悬液以1 000 r/min转速,离心5 min,弃上清液,加入2 ml完全培养基重悬,并按1∶2的比例均匀接种于两个培养皿,加入8 ml完全培养基水平摇匀后置于恒温细胞培养箱内培养,取对数生长期的细胞进行后续细胞毒性实验。
-
将实验细胞单独用药组根据药物种类与浓度不同,分为hcGEM单药组(hcGEM: 0.015 625、0.031 25、0.062 5、0.125、0.25、0.5、1、2、4 μmol/L),Era单药组(Era:0.25、0.5、1、2、4、8、16、32、64 μmol/L),SASP单药组(SASP: 6.25、12.5、25、50、100、200、400、800、1600 μmol/L)以及Art单药组(Art:0.5、1、2、4、8、16、32、64、128 μmol/L),每种药物都按2倍比率设置浓度梯度,每组各9个浓度;联合用药组根据联合药物组成与比例不同分为hcGEM-Era联合用药组(包括4∶1、1∶1、1∶4和1∶16 联合组)、hcGEM-SASP联合用药组(包括4∶1、1∶1、1∶20和1∶400联合组)、hcGEM-Art联合用药组(包括 2∶1、1∶1、1∶4和1∶16联合组)。每个联合用药组中,hcGEM的浓度均为相同的浓度梯度(hcGEM: 0.015 625、0.031 25、0.062 5、0.125、0.25、0.5、1、2、4 μmol/L),并与恒定比例的联合药物共同作用于细胞。hcGEM-Era 1∶16联合组的药物浓度梯度为0.015 625 μmol/L hcGEM+0.25 μmol/L Era、0.031 25 μmol/L hcGEM+0.5 μmol/L Era、0.062 5 μmol/L hcGEM+1 μmol/L Era、0.125 μmol/L hcGEM+2 μmol/L Era、0.25 μmol/L hcGEM+4 μmol/L Era、0.5 μmol/L hcGEM+8 μmol/L Era、1 μmol/L hcGEM+16 μmol/L Era、2 μmol/L hcGEM+32 μmol/L Era、4 μmol/L hcGEM+64 μmol/L Era。
-
取对数生长的PANC-1细胞接种于96孔板,密度为5 000个细胞/孔。待细胞贴壁24 h后,弃上清液,每3个复孔加入100 μl不同浓度的每种药物,另设空白对照组与药物未处理组各3个复孔。药物与细胞共培养48 h后,每孔加入含10% CCK-8的DMEM培养基。于恒温培养2 h后,用酶标仪测量450 nm波长下各孔的吸光度值(A值)。使用GraphPad Prism 8.0.2软件对数据进行非线性回归分析,得到各组药物作用于细胞的半数抑制浓度(IC50) 以及细胞增殖抑制率,具体公式为:抑制率=[1−(A用药组−A空白组)/(A未用药组−A空白组)]×100%。
-
根据单独用药组与联合用药组的摩尔浓度与细胞增殖抑制率,使用CompuSyn软件计算联合指数(CI),并依据CI值判断药物的联合效果: CI>1表示拮抗作用;CI=1表示相加作用;CI<1表示协同作用,且协同抑制效果随CI值的减小而增强。
-
实验结果以(
$\bar x $ ±s)表示,采用GraphPad Prism 8.0.2软件进行数据的处理分析,多组间比较采用单因素方差分析,两组间比较采用t检验。P<0.05表示组间差异有统计学意义。 -
如图1所示,hcGEM、Era、SASP、Art单独作用于PANC-1细胞时可明显抑制细胞的增殖,并且这种抑制作用呈现剂量依赖性。hcGEM、Era、SASP、Art作用于PANC-1细胞的IC50值分别为0.175 7、1.884、195.1、23.13 μmol/L。hcGEM单独用药时,小于0.015 625 μmol/L的剂量对PANC-1细胞的生长几乎无抑制作用,存活率>95%(P>0.05);剂量大于2 μmol/L时,细胞的活性受到显著抑制,存活率<15%(P<0.05)。Era单独用药时,大于16 μmol/L 的剂量能显著抑制PANC-1细胞的生长,存活率<10%(P<0.05),而小于0.25 μmol/L的剂量会使80%以上的细胞存活。SASP单独用药时,大于800 μmol/L的剂量能抑制PANC-1细胞的生长,存活率<15%(P<0.05),而小于12.5 μmol/L的剂量仅轻微抑制PANC-1细胞存活,存活率>85%(P>0.05)。Art单独用药时,大于128 μmol/L的剂量能显著抑制PANC-1细胞的生长,存活率<40%(P<0.05),而小于0.5 μmol/L时,仅能轻微抑制细胞增殖,存活率>85%(P>0.05)。
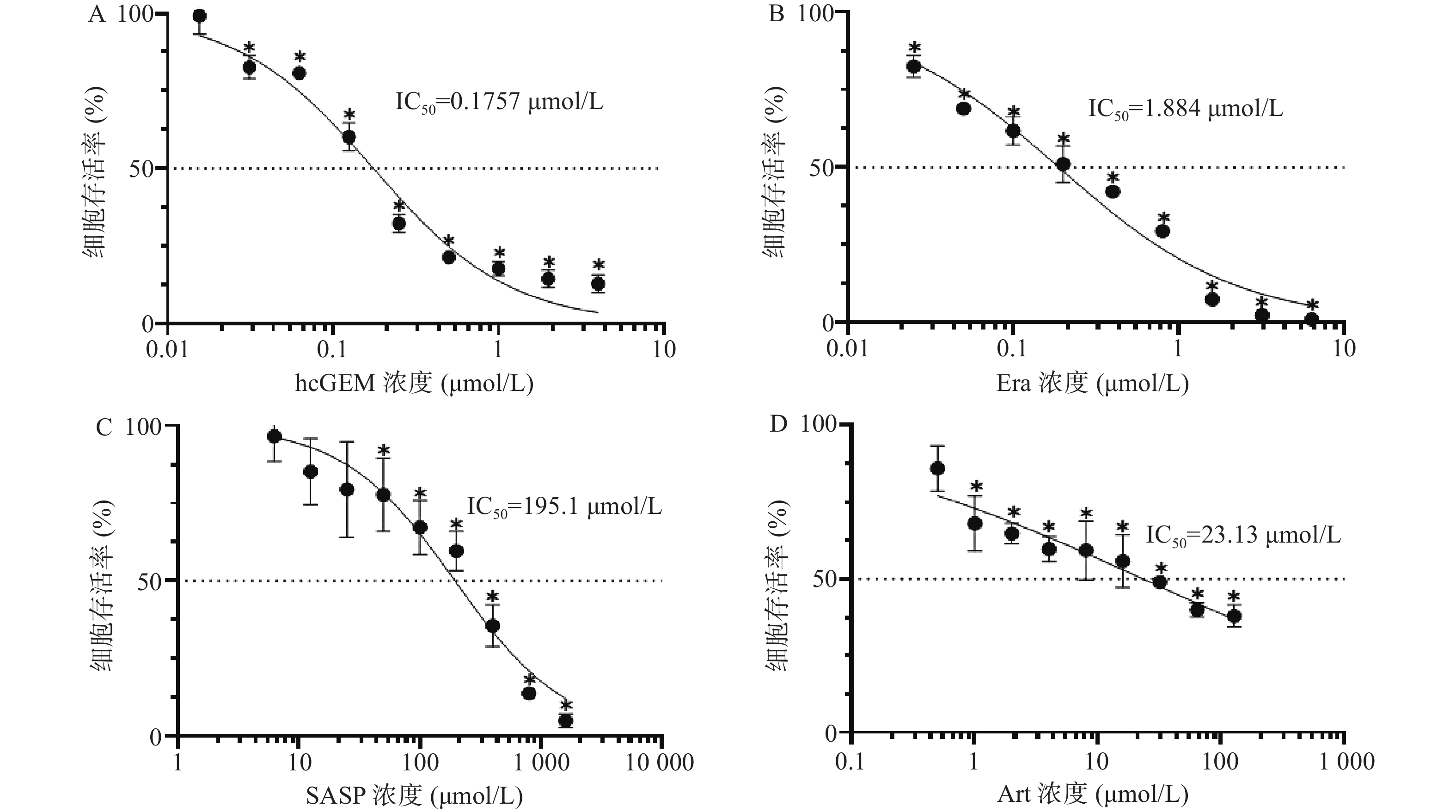
图 1 不同浓度的三种铁死亡诱导剂对胰腺癌PANC-1细胞存活率的影响
-
在研究单药对PANC-1细胞抑制效果的基础上,我们进一步探究了hcGEM-Era 4∶1、1∶1、1∶4和1∶16 联合用药,hcGEM-SASP 4∶1、1∶1、1∶20和1∶400联合用药,以及hcGEM-Art 2∶1、1∶1、1∶4和1∶16联合用药,对PANC-1细胞的抑制效果。如图2所示, hcGEM-Era、hcGEM-SASP以及hcGEM-Art联合用药组的抑制效果随着浓度的增加而提高。其中,hcGEM-Era 4∶1、1∶4、1∶16联合组,以及hcGEM-SASP 1∶400联合组对PANC-1细胞抑制效果在所有浓度梯度范围均优于hcGEM组(P<0.05)。此外,hcGEM-Era 4∶1、1∶4、1∶16联合用药组,hcGEM-SASP 1∶20、1∶400联合组,和hcGEM-Art 1∶16联合组的IC50值均小于0.1757 μmol/L(hcGEM单药组的IC50),说明上述联合用药能在一定程度上提高对PANC-1细胞的抑制效果。不同联合用药组hcGEM的IC50值见表1。
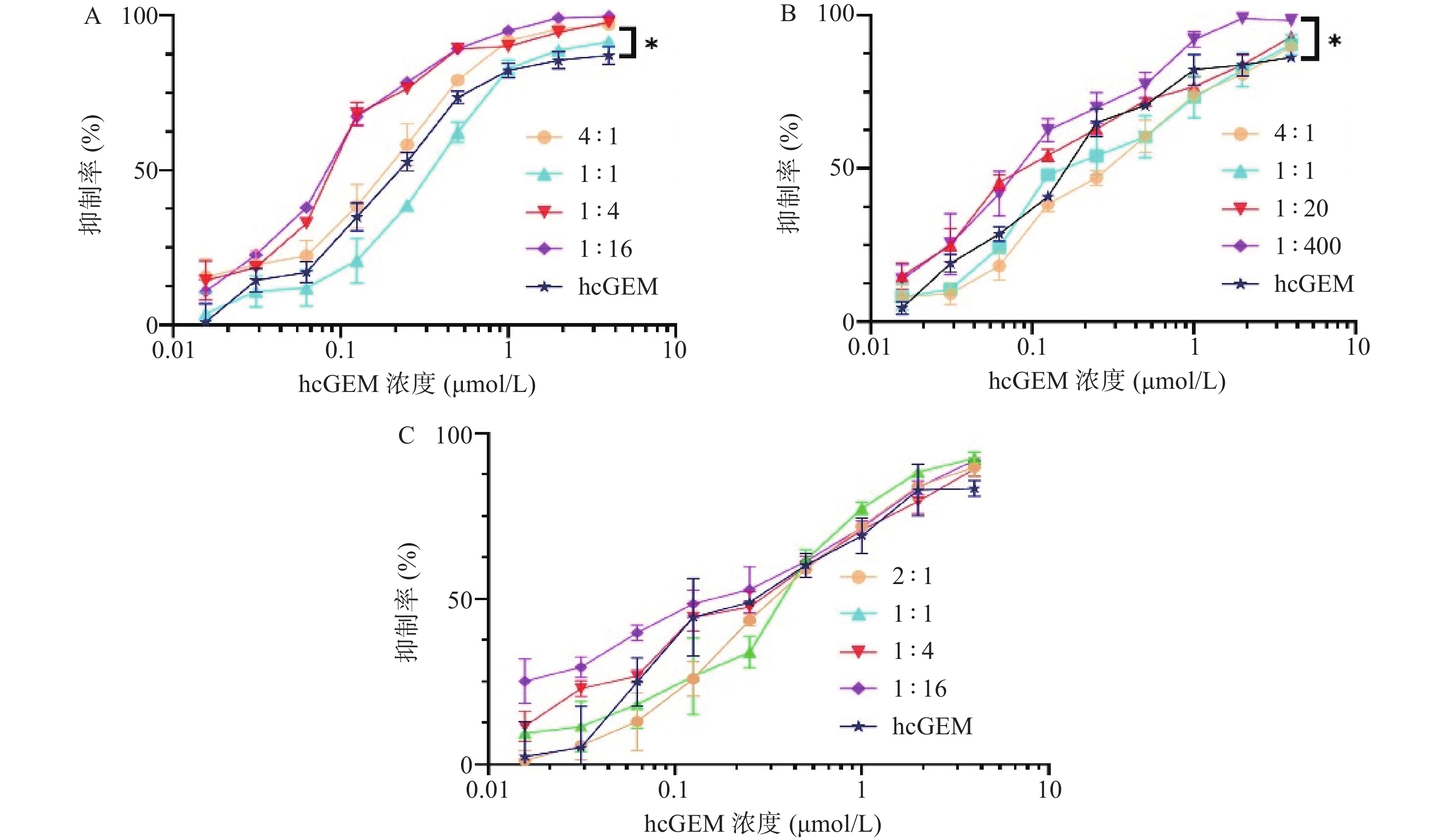
图 2 三种铁死亡诱导剂分别联用hcGEM对胰腺癌PANC-1细胞存活率的影响
表 1 hcGEM联合三种铁死亡诱导剂对胰腺癌PANC-1细胞作用时的IC50值
组别 摩尔浓度比 IC50(μmol/L,hcGEM) hcGEM-Era联合组 1∶0.25 0.140 3±0.009 1 1∶1 0.340 2±0.018 3 1∶4 0.091 3±0.005 1 1∶16 0.083 3±0.002 5 hcGEM-SASP联合组 1∶0.25 0.297 5±0.016 1 1∶1 0.240 2±0.021 2 1∶20 0.120 8±0.008 9 1∶400 0.092 6±0.006 7 hcGEM-Art联合组 1∶0.5 0.366 4±0.018 7 1∶1 0.344 4±0.026 3 1∶4 0.249 3±0.015 7 1∶16 0.154 6±0.013 5 -
为了研究hcGEM分别与Era、SASP、Art联合用药,对PANC-1细胞是否具有协同抑制作用,我们分别设计了hcGEM与三种铁死亡诱导剂的4种不同比例的联用方案,来探讨不同联合用药组对PANC-1细胞的协同抑制效果。CompuSyn软件分析结果显示,hcGEM-Era 4∶1或1∶4联用组在所有药物浓度下均能对PANC-1细胞产生良好的协同抑制效果(CI<1),且4∶1联用组的协同抑制效果略优于1∶4联用组。对于hcGEM-Era 1∶16联用组,CI值随着联合药物浓度的增加而减小,说明该比例下两药协同抑制效果随着浓度增大有所增强,而hcGEM-Era 1∶1联用组的CI值,仅除1 μmol/LhcGEM+1 μmol/L Era、0.031 25 μmol/L hcGEM+0.031 25 μmol/L Era联用组的CI<1外,其余浓度组CI>1,说明hcGEM-Era 1∶1联用组对PANC-1细胞几乎无协同抑制效果(图3A)。
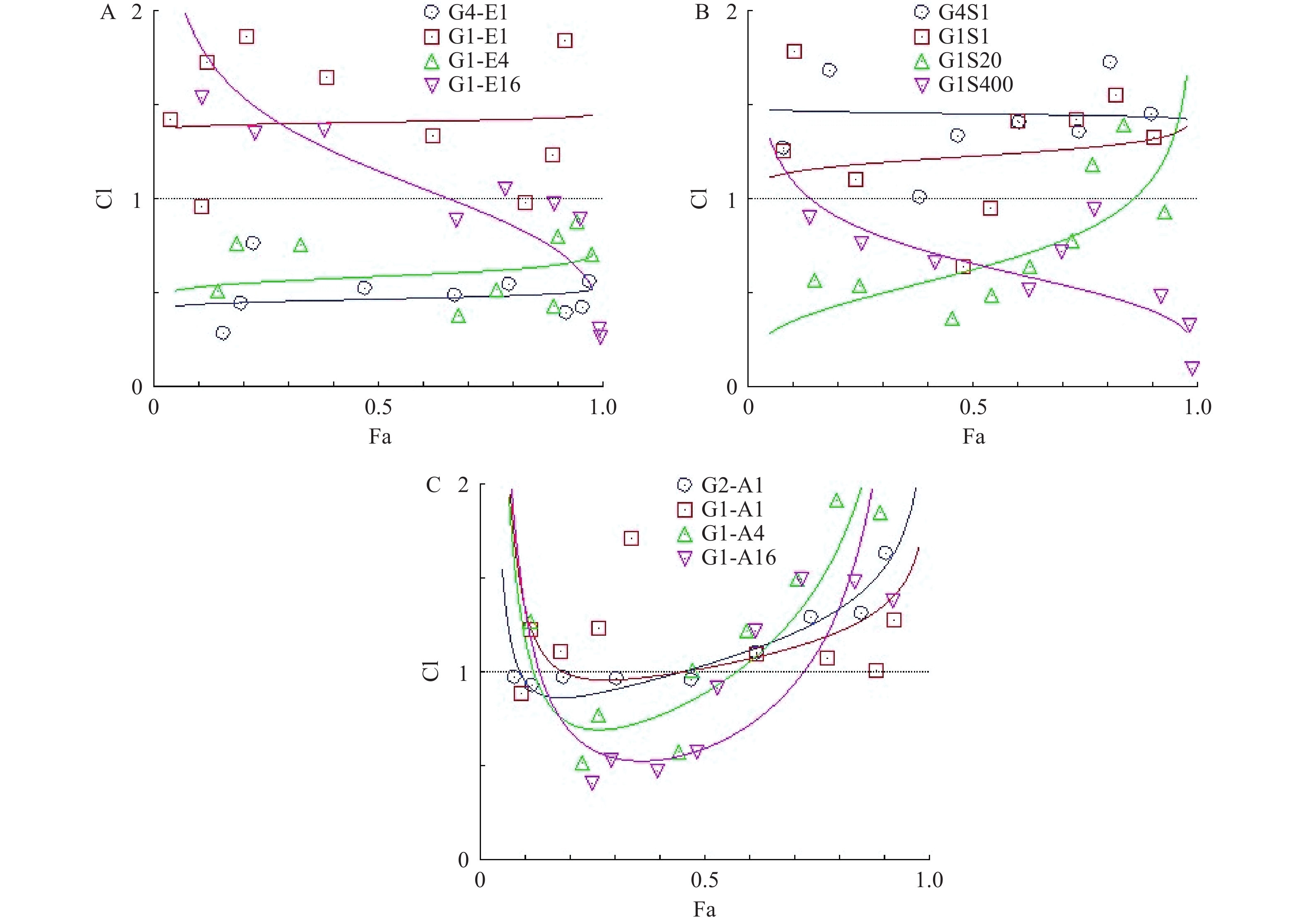
图 3 三种铁死亡诱导剂分别联用hcGEM对胰腺癌PANC-1细胞的效应-联合指数曲线图
对于hcGEM与SASP联合用药组对PANC-1细胞的协同抑制效果,CompuSyn分析结果提示,hcGEM-SASP 4∶1联用对PANC-1细胞没有协同作用(CI>1),而hcGEM-SASP 1∶1联用组除了0.25 μmol/L hcGEM+0.25 μmol/L SASP、0.125 μmol/L hcGEM+0.125 μmol/LSASP联用组CI<1外,其余联用浓度组的CI>1,表明hcGEM-SASP 1∶1联合用药对PANC-1细胞几乎无协同抑制作用。hcGEM-SASP 1∶20联合用药对PANC-1细胞,除了1 μmol/LhcGEM+20 μmol/L SASP、2 μmol/L hcGEM+40 μmol/LSASP联用组CI>1外,其余浓度组CI<1,提示hcGEM-SASP 1∶20联合用药对PANC-1细胞主要是协同抑制效果,且协同效果随着浓度的增加而减弱。hcGEM-SASP 1∶400联合用药对PANC-1细胞有良好的协同抑制效果(CI<1),且协同效果随着浓度的增加而增强(图3B)。
对于hcGEM与Art联合用药组对PANC-1细胞的协同抑制效果,CompuSyn结果提示,hcGEM-Art 2∶1联用组在0.031 25 ~0.25 μmol/L hcGEM联合浓度范围内CI <1,且接近1,其余联合浓度组的CI>1,表明hcGEM-Art 2∶1联合用药对PANC-1细胞几乎无协同抑制作用。hcGEM-Art 1∶1联用组CI>1,表明hcGEM-Art 1∶1联合用药对PANC-1细胞无协同抑制作用。hcGEM-Art 1∶4联用组仅在0.031 25~0.125 μmol/L hcGEM联合浓度范围内CI<1,其余联合浓度范围内CI>1,表明hcGEM-Art 1∶4联合用药对PANC-1细胞仅在0.031 25~0.125 μmol/L hcGEM联合浓度范围内存在协同抑制作用。 hcGEM-Art 1∶16联用组在0.015 625~0.25 μmol/L hcGEM联合浓度范围内CI <1,其余联合浓度范围内CI>1,表明hcGEM-Art 1∶16联用组在0.015 625~0.25 μmol/L hcGEM联合浓度范围内对PANC-1细胞有协同抑制作用 (图3C)。
-
目前,胰腺癌的一线治疗标准为吉西他滨联合白蛋白结合型紫杉醇或者FOLFIRONOX组合(5-氟尿嘧啶、亚叶酸钙、伊立替康和奥沙利铂)[10]。吉西他滨在一定程度上能提高患者的生存率,但耐药性的出现以及毒副作用限制了临床治疗效果,故临床上常将其与其他化疗药联合使用来提高其疗效[11]。近来大量研究证实,诱导癌细胞铁死亡可能对包括胰腺癌在内的多种类型癌症有效[12]。自2003年发现小分子铁死亡诱导剂埃拉斯汀至今,研究人员已发现多种铁死亡诱导剂,如RSL3、Sorafeni、SASP、Art等[13]。本研究选取了三种铁死亡诱导剂Era、SASP以及Art,首先研究了三种铁死亡诱导剂单独应用对PANC-1细胞的增殖抑制作用。通过CCK-8法检测了三种铁死亡诱导剂的细胞毒性作用,我们发现Era对PANC-1细胞的IC50最小,而SASP的IC50最大,表明在这三种铁死亡诱导剂中,Era对PANC-1细胞的抑制作用最强,而SASP最弱。三种铁死亡诱导剂单独使用对PANC-1细胞的增殖抑制作用呈现剂量依赖性,即浓度越大抑制效果越强。
为了进一步探究三种铁死亡诱导剂Era、SASP、Art与hcGEM联用是否对胰腺癌PANC-1细胞具有协同抑制作用,本研究设计了不同比例的联合用药组来探索最佳的联合协同方案。本次研究结果显示,当三种铁死亡诱导剂分别与hcGEM联合使用时, hcGEM-Era 4∶1或1∶4联用组,hcGEM-SASP 1∶400联用组对PANC-1细胞具有良好的协同抑制效果。hcGEM-Art 1∶4或1∶16联用组仅在一定浓度范围内对PANC-1细胞有协同抑制效果。胰腺导管腺癌患者中,90%存在KRAS突变, 而致癌KRAS将胰腺导管腺癌细胞重新编程为高度抗凋亡的状态。由于KRAS信号突变的副产物是生成大量的ROS,为了上调抗氧化能力,胰腺导管腺癌转而增强葡萄糖和谷氨酰胺代谢途径,使细胞对ROS诱导的、铁依赖性非凋亡的铁死亡模式敏感[13]。激活铁死亡可有效阻止肿瘤进展,增强化疗、放疗和免疫治疗的效果[14]。在治疗胰腺癌的过程中,吉西他滨通过NF-κB信号途径诱导内源性活性氧的产生,进而导致Nrf2信号通路的激活以及细胞内谷胱甘肽增加,最终使胰腺癌细胞对吉西他滨不敏感[15]。埃拉斯汀可通过抑制胱氨酸-谷氨酸逆向转运蛋白的活性来减少胱氨酸进入肿瘤细胞,从而降低胞内谷胱甘肽的合成,抑制线粒体的电压依赖性阴离子通道等多种途径促进线粒体代谢紊乱、活性氧类物质以及脂质过氧化物的大量积累,增强吉西他滨对胰腺癌细胞的杀伤效果[16]。柳氮磺胺吡啶可通过抑制胱氨酸-谷氨酸逆向转运蛋白的功能促进胰腺癌细胞的铁死亡,逆转癌细胞的耐药性 [17]。青蒿琥酯通过诱导铁依赖性氧化损伤促进胰腺癌细胞的死亡,并且这种脂质过氧化性细胞死亡可被铁死亡抑制剂阻断[18]。本研究中三种铁死亡诱导剂联合hcGEM对PANC-1细胞的协同抑制的效果可能与这三种铁死亡诱导剂引起的氧化应激反应的强弱有关,具体相关机制还有待进一步实验。
综上所述,本研究发现三种铁死亡诱导剂分别与hcGEM联合应用时存在对PANC-1细胞的协同抑制联合方案。hcGEM-Era 4∶1或1∶4联合用药以及hcGEM-SASP 1∶400联合用药对PANC-1细胞的协同抑制效果良好,而hcGEM-Art 1∶4或1∶16联合用药仅一定浓度范围能对PANC-1细胞产生协同抑制效果。
Inhibitory effects of gemcitabine hydrochloride combined with ferroptosis inducers on the proliferation of PANC-1 cells
-
摘要:
目的 研究三种铁死亡诱导剂埃拉斯汀(Era)、柳氮磺胺吡啶(SASP)和青蒿琥酯(Art)与盐酸吉西他滨(hcGEM)单独或联合应用对人胰腺癌PANC-1细胞的抑制作用。 方法 采用CCK-8法检测不同浓度的Era、SASP、Art单独或联合hcGEM对PANC-1细胞的增殖抑制作用,并根据联合指数(CI) 判断三种铁死亡诱导剂联合hcGEM对 PANC-1细胞是否具有协同抑制作用。 结果 hcGEM与三种铁死亡诱导剂单独或联合应用均能显著抑制PANC-1细胞活性,并且这种抑制作用随着浓度增大而增强。hcGEM-Era 4∶1或1∶4联用组、hcGEM-SASP 1∶400联用组 CI 值小于1。hcGEM-Art 1∶4或1∶16联用组仅在一定浓度范围内CI值小于1。 结论 hcGEM与三种铁死亡诱导剂单独或联合应用对PANC-1细胞的增殖抑制作用呈现剂量依赖性。hcGEM与三种铁死亡诱导剂联用能协同抑制PANC-1细胞的增殖。 Abstract:Objective To study the effects of three ferroptosis inducers Erastin (Era), sulfasalazine (SASP) and artesunate (Art) alone or combined with gemcitabine hydrochloride (hcGEM) on the proliferation inhibition of Human pancreatic cell line PANC -1. Methods The CCK-8 method was used to detect the inhibitory effects of different concentrations of Era, SASP and Art alone or combined with hcGEM on the proliferation of PANC-1, and the combination index (CI) was used to judge whether three ferroptosis inducers combined with hcGEM had synergistic inhibitory effect on PANC-1. Results The three ferroptosis inducers and hcGEM alone or in combination could significantly inhibit the activity of PANC-1. The inhibitory effects were enhanced with the concentration increasing. The CI values of hcGEM-Era 4∶1 or 1∶4 combination group and hcGEM-SASP 1∶400 combination group were less than 1.The CI values of hcGEM-Art 1∶4 or 1∶16 combination group were less than 1 only within a certain concentration range. Conclusion The inhibitory effects of the three ferroptosis inducers and hcGEM alone or in combination were dose-dependent. The combination of hcGEM and three ferroptosis inducers could synergistically inhibit the proliferation of PANC-1. -
多囊卵巢综合征(polycystic ovary syndrome,PCOS)是育龄期女性最常见的一种生殖内分泌疾病,其特征以生化或临床高雄激素血症、无排卵和卵巢多囊样改变等为主要表现[1-2]。流行病学显示PCOS在全世界范围内的总体发病率约为6%~20%[3],而由PCOS引发的排卵障碍所致的不孕症占40%[4]。PCOS不仅影响女性的正常生殖功能,还会导致女性发生代谢系统方面的障碍,如高胰岛素血症、胰岛素抵抗,增加女性继发糖尿病、心血管疾病及子宫内膜癌的风险,对女性健康造成严重不良影响[2]。肥胖是PCOS发生的重要危险因素之一[5]。研究数据表明,28.3%的超重或肥胖的女性患有PCOS[6],多达42%的PCOS患者超重或肥胖[7]。肥胖对PCOS的发展和进展产生显著影响,研究发现,脂肪细胞主要通过分泌脂肪因子,如IL-1、IL-6、瘦素、脂联素等,作用于相应的靶器官、靶组织、靶细胞,如卵巢、肾上腺等,刺激机体产生较多的雄激素,而雄激素又可通过抑制肾上腺素受体等导致体内脂肪分解减少,脂肪大量堆积在体内,导致体内高雄激素水平与肥胖之间形成恶性循环,严重影响PCOS患者的健康状况[3]。与普通女性相比,PCOS患者具有更高的肥胖倾向,且更容易出现腹部脂肪堆积[8];而这种由于腹部脂肪堆积造成的中心性肥胖反过来又可加重PCOS患者的临床或生化表现,导致胰岛素抵抗、高雄激素血症、生殖功能异常等[9]。临床试验结果显示,若患者体质量减轻初始体质量的5%,其体内激素水平、血糖水平得以改善,同时,月经周期和排卵情况趋于正常化,这表明体质量减轻可增加患者排卵和妊娠的可能性[10]。
青蒿素(artemisinin,ART)是一种天然倍半萜内酯化合物,最初由2015年诺贝尔生理学或医学奖获得者屠呦呦从青蒿植物中提取出来并广泛用于抗疟疾治疗[11];青蒿素还用于抗癌、抗炎药物等[12]。近年研究发现,青蒿素及其衍生物还具有预防肥胖的功效:在啮齿动物模型中,青蒿素及其衍生物通过调节p38MAPK/ATF2轴和Akt/mTOR途径等在脂肪生成过程中诱导脂肪细胞褐变,从而预防肥胖并改善肥胖相关的代谢紊乱[13]。Lee等[14]和Jang[15]体外实验数据表明,青蒿素及其衍生物可通过PPARγ途径抑制脂肪生成和脂肪因子的表达。本研究通过网络药理学方法和分子对接方法分析预测青蒿素可用于治疗PCOS的潜在靶点,旨在为深入研究其治疗的作用机制提供参考。
1. 材料和方法
1.1 青蒿素及PCOS靶点预测
通过Pubchem数据库[16](
https://pubchem.ncbi.nlm.nih.gov/ )获得天然产物青蒿素的SMILES号,并将其输入Swiss TargetPrediction数据库[17](http://www.swisstargetprediction.ch/ )进行靶点预测,导出分析结果并保存;结合PharmMapper数据库[18](http://www.lilab-ecust.cn/pharmmapper/ )预测的靶点,二者共同作为青蒿素的药物靶点,并将靶点导入Uniprot数据库[19](https://www.uniprot.org/)进行靶点蛋白与基因名称转换。以“polycystic ovary syndrome”作为关键词检索,通过DisGeNET数据库[20](https://www.disgenet.org/ )、GeneCard数据库[21](https://www.genecards.org/ )进行疾病靶点预测。将搜集的青蒿素靶点和PCOS靶点分别导入Venny在线作图软件(https://bioinfogp.cnb.csic.es/tools/venny/ )绘制韦恩图,从而得到二者的共同靶点。1.2 PPI网络构建及核心靶点筛选
将共同靶点导入STRING数据库[22](
https://cn.string-db.org/ ),物种选择“homo sapiens”,最低相互作用分数设置为“0.9”,隐藏游离点,其他保持默认设置,得到蛋白相互作用网络图(protein-protein interaction,PPI),将PPI网络图导入Cytoscape 3.9.1软件[23],进行核心靶点筛选。1.3 富集分析
将共同靶点导入DAVID数据库[24](
https://david.ncifcrf.gov/ ),分别进行基因本体(gene ontology,GO)功能、京都基因与基因组百科全书(kyotoencyclopodia of genes and genomes,KEGG)通路分析,其中GO功能富集内容从分子功能(molecular function,MF)、生物学过程(biological process,BP)、细胞组分(cellular component,CC)三部分进行逐一分析,并利用微生信在线作图软件(http://www.bioinformatics.com.cn/ )将分析结果进行可视化。1.4 药物-疾病-靶点-通路网络图构建
将所获得的青蒿素、PCOS作用靶点及信号通路分别导入Cytoscape软件构建药物-疾病-靶点-通路网络图。
1.5 分子对接
从Pubchem数据库中下载青蒿素的2D结构,在RCSB PDB数据库[25](
https://www.rcsb.org/ )中下载核心靶蛋白结构。利用Chem3D软件对青蒿素的2D结构进行转化,用Pymol软件对核心靶蛋白结构进行初步处理,再用Auto Dock Tools软件做进一步加氢等处理,并将处理的核心靶蛋白保存为“pdbqt”格式进行分子对接,最后利用Pymol软件对分子对接结果进行可视化处理[26-27]。2. 结果
2.1 青蒿素及PCOS作用靶点
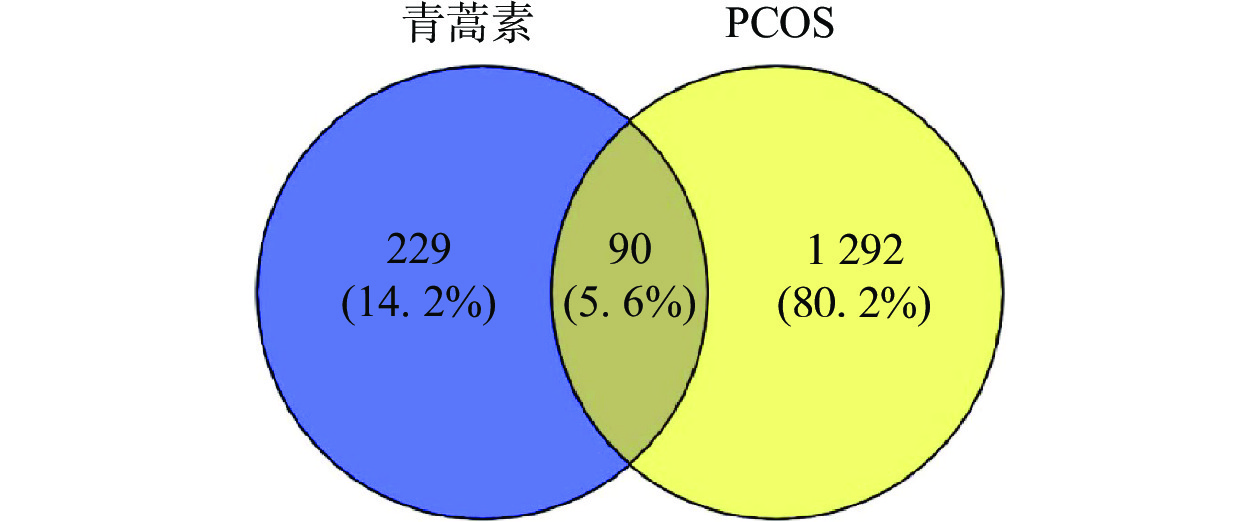
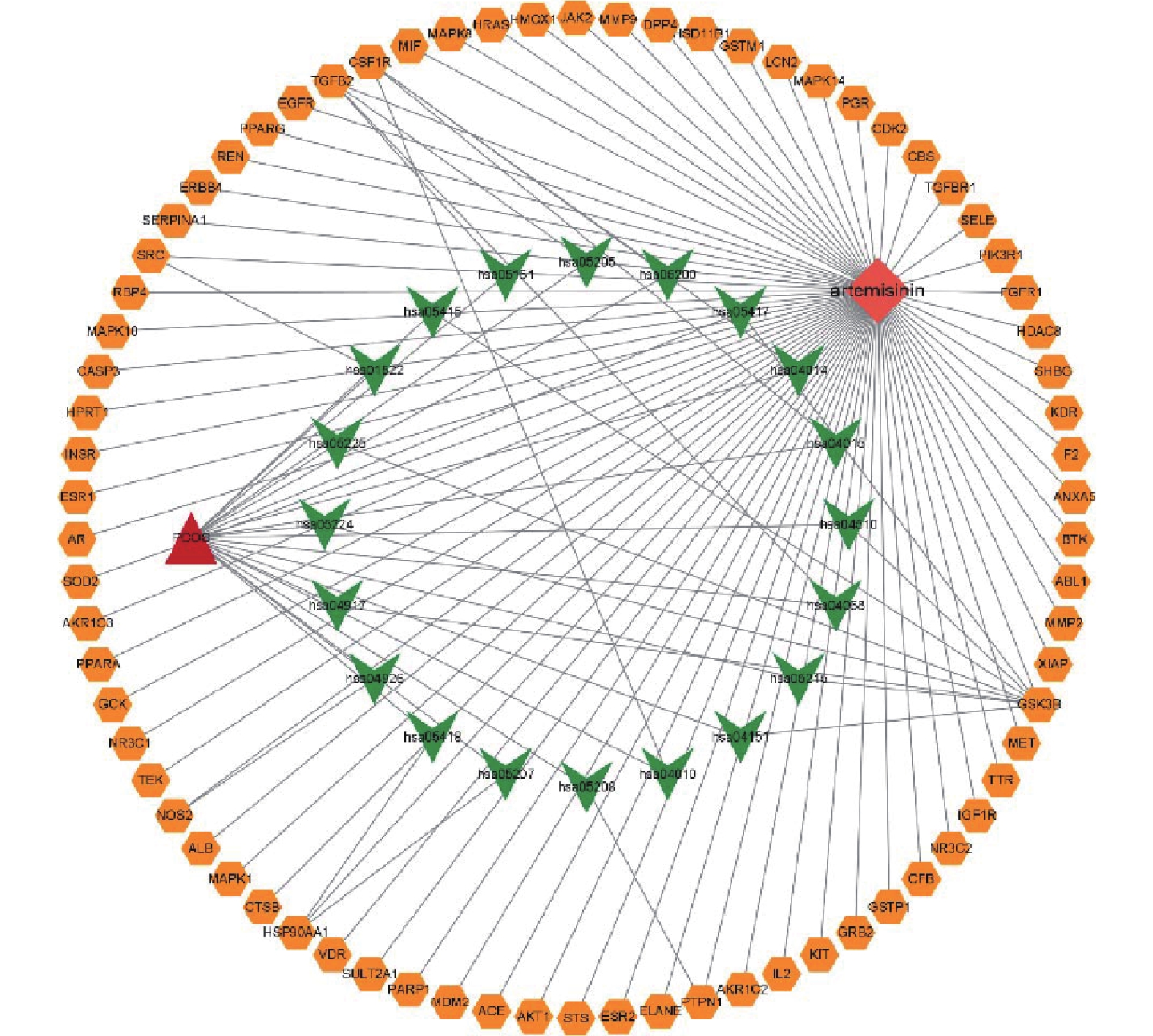
通过数据库检索共得到青蒿素潜在作用靶点229个,PCOS疾病靶点1292个。利用在线作图软件将青蒿素作用靶点与PCOS疾病靶点进行韦恩图分析,得到二者的交集靶点90个,如图1所示。
2.2 PPI网络构建及核心靶点筛选
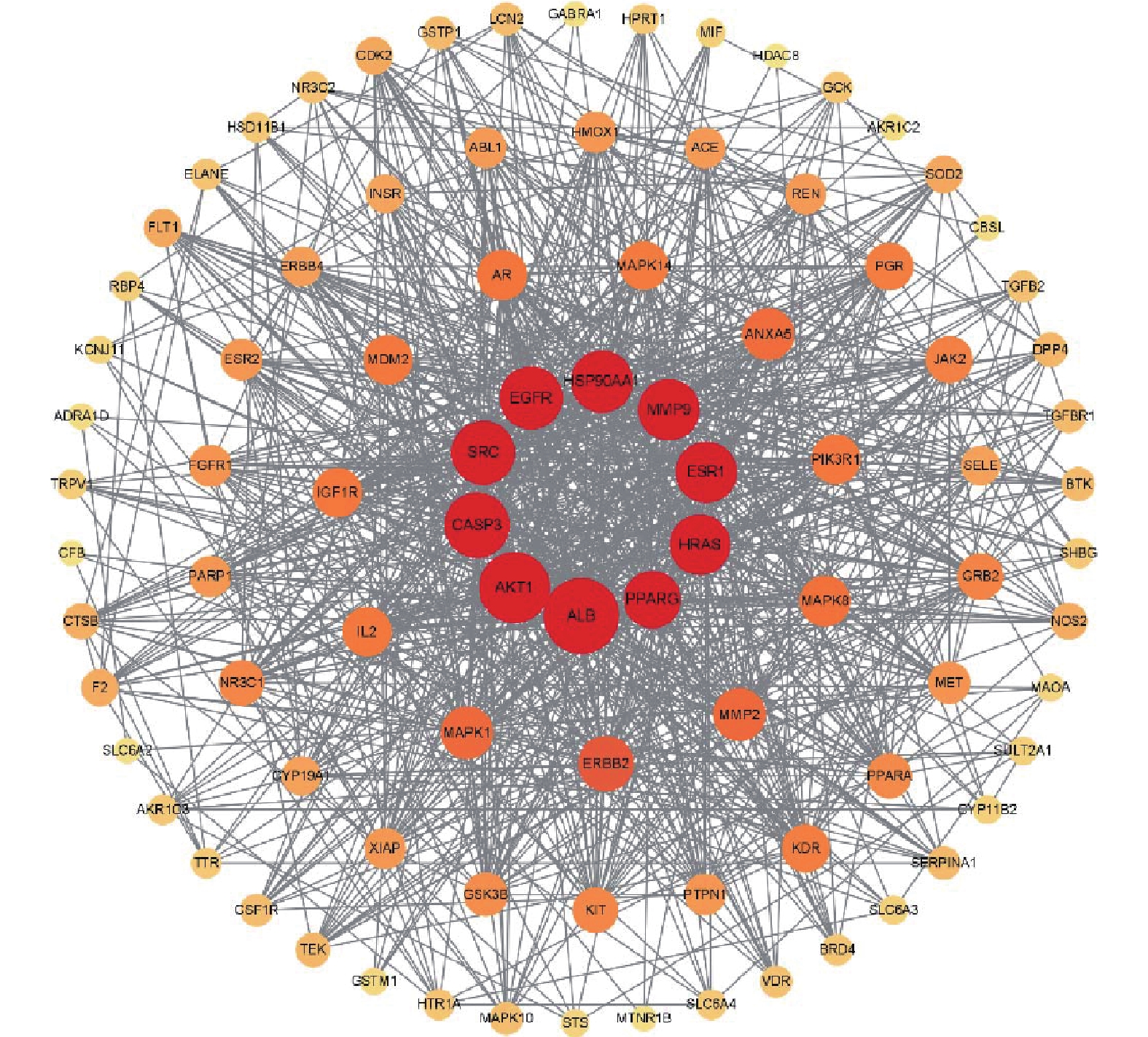
将交集靶点导入String数据库,绘制PPI网络关系图,如图2所示,其中包括网络节点90个,边235条。将共同靶点导入Cytoscape 软件进行核心靶蛋白筛选。如图3所示。综合节点度值及本研究相关度排名靠前的分别为AKT1、ESR1、MMP9、PPARγ、MMP2(见表1)。
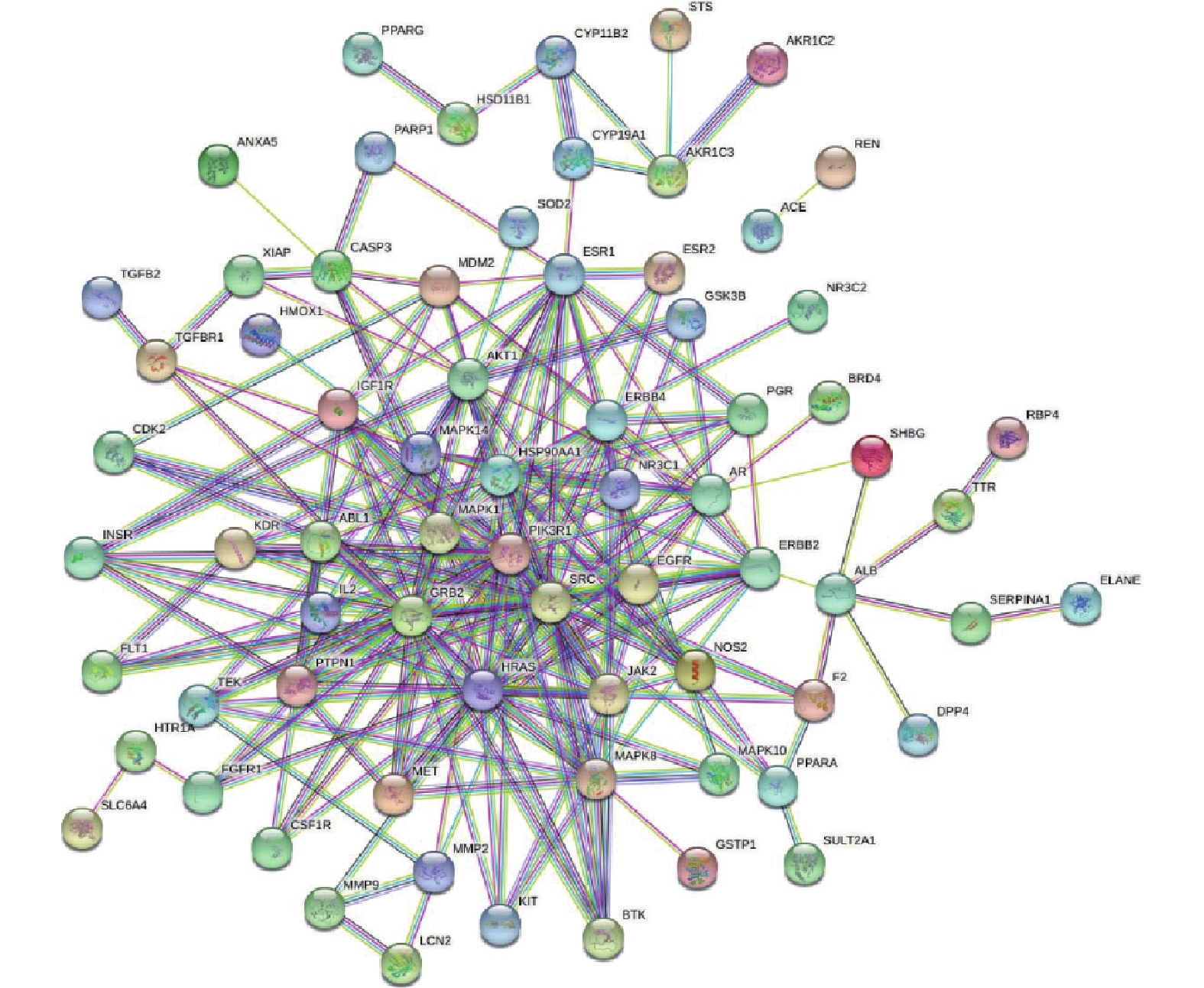 图 2 蛋白互作网络图注:图中节点代表蛋白质,其中红色节点表示查询蛋白质,其他颜色节点表示与查询蛋白只有相互作用的其他蛋白质,空白节点表示未知3D结构的蛋白质,填充节点表示已知3D结构,连线代表蛋白与蛋白之间的相互作用关系表 1 青蒿素作用于PCOS的核心靶点
图 2 蛋白互作网络图注:图中节点代表蛋白质,其中红色节点表示查询蛋白质,其他颜色节点表示与查询蛋白只有相互作用的其他蛋白质,空白节点表示未知3D结构的蛋白质,填充节点表示已知3D结构,连线代表蛋白与蛋白之间的相互作用关系表 1 青蒿素作用于PCOS的核心靶点基因名称 节点度值 排名 ALB 66 1 AKT1 60 2 CASP3 53 3 SRC 51 4 EGFR 50 5 HSP90AA1 49 6 MMP9 48 7 ESR1 48 8 HRAS 47 9 PPARγ 43 10 ERBB2 41 11 MMP2 37 12 2.3 富集分析
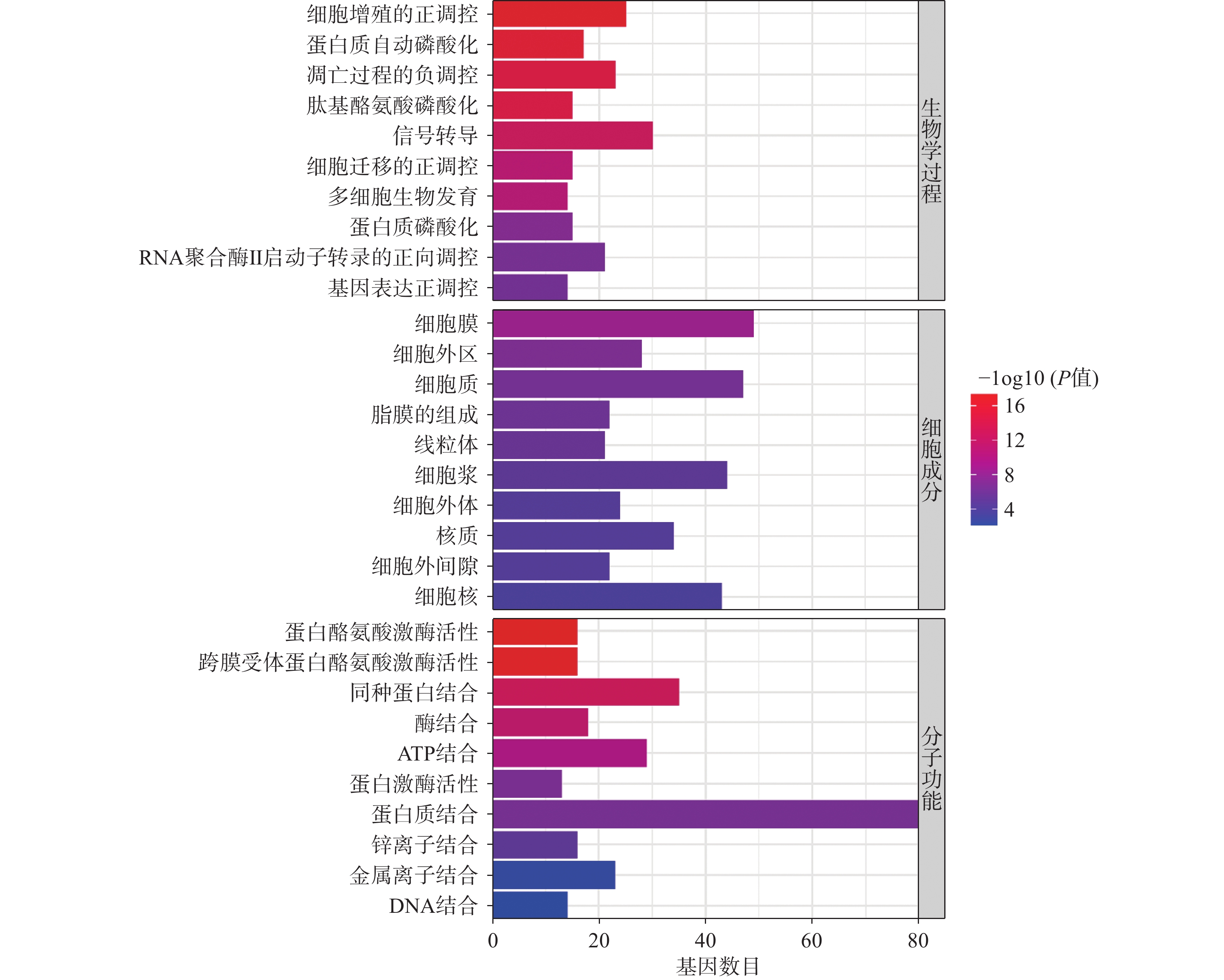
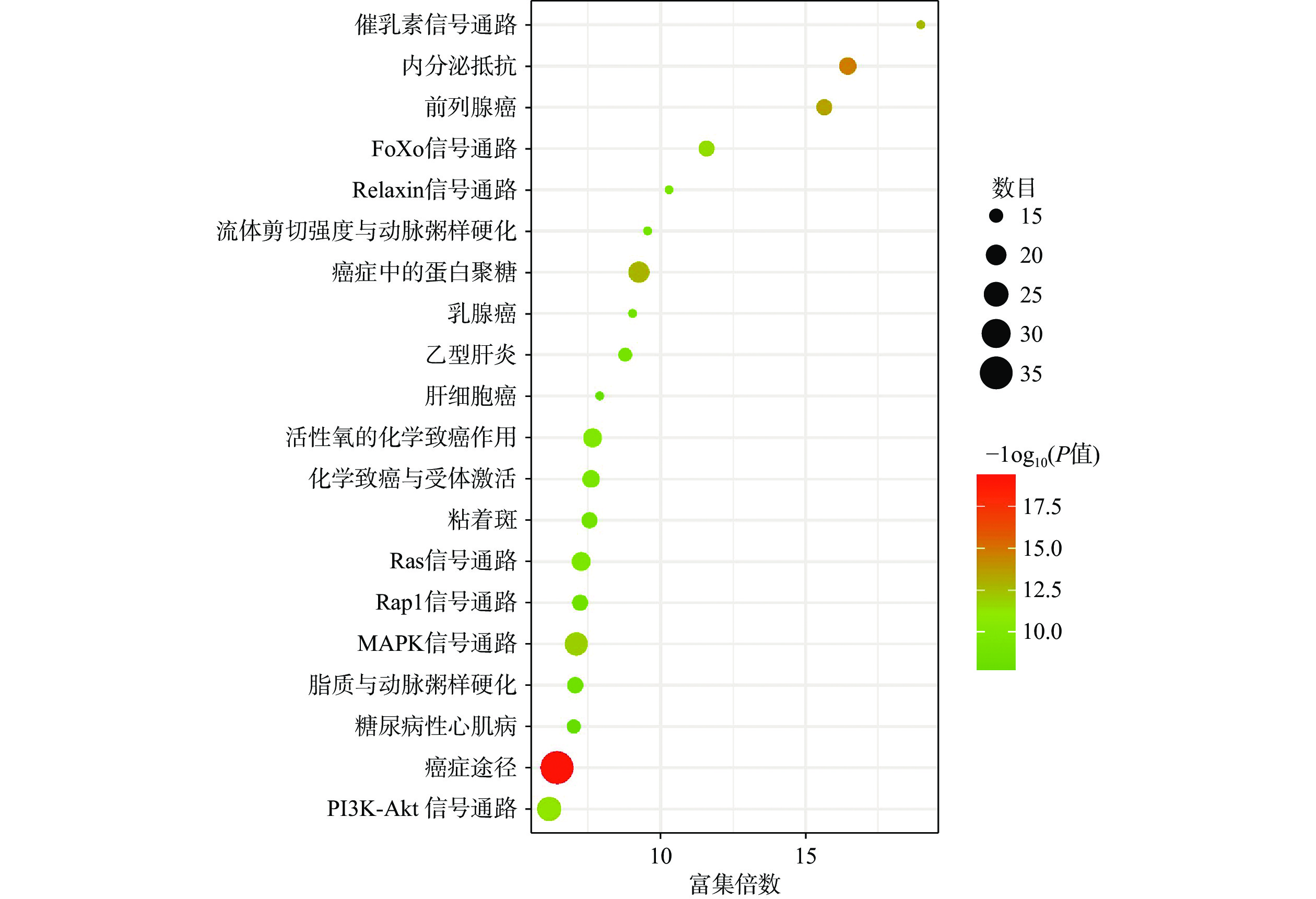
将得到的90个交集靶点导入DAVID数据库进行GO富集分析,富集结果分别根据基因富集程度进行排序,其中BP前10个条目主要涉及细胞增殖调控、蛋白质磷酸化和RNA聚合酶Ⅱ启动子转录的正调控等生物学过程,MF前10个条目主要与蛋白酪氨酸激酶活性、蛋白激酶活性、蛋白结合和酶结合等分子功能有关,CC前10个条目主要在细胞膜、胞质和胞核等部位富集,如图4所示。KEGG富集分析共筛选到162条信号通路,根据基因富集程度排序,前20个条目主要涉及PI3K/Akt、MAPK、Ras、内分泌抵抗等信号通路,如图5所示。
2.4 药物-疾病-靶点-通路网络图构建
将相关靶点及通路文件导入Cytoscape 3.9.1软件,得到药物、疾病、靶点和通路之间的关系图(见图6)。
2.5 分子对接
分子对接结果显示,青蒿素与核心靶蛋白AKT1、MMP9、ESR1、PPARγ、MMP2之间均存在结合位点。青蒿素与核心靶蛋白的最低结合能分数见表2,结合能越低表示结合活性越高,化合物越容易与该靶点结合。其中青蒿素与核心靶蛋白之间的氢键连接可视化情况如图7所示。
表 2 青蒿素与核心靶点分子对接结果化合物 核心靶点 最低结合能(kJ/mol) 结合位点 青蒿素 MMP9 −8.2 ARG-143 AKT1 −7.9 HIS-152 ESR1 −7.9 THR-460 PPARγ −7.7 PRO-426、GLN-430、
LEU-431、PHE-432MMP2 −6.4 HIS-190 3. 讨论
PCOS主要通过卵巢病变、机体内分泌紊乱等方式影响女性的生育能力[28]。目前关于PCOS的发病机制和病因尚未具体阐明,多认为是遗传和环境因素相互作用的结果;由于病因机制不明,临床治疗尚无统一方案,多采用对症治疗,如基础生活方式调整,通过控制饮食、增加体育运动以降低体质量和缩小腰围,增加机体胰岛素敏感性,降低胰岛素及雄激素水平,同时辅以相应的药物治疗,以减轻症状。
本研究基于临床发现,PCOS患者多伴有肥胖表现,遂以肥胖与PCOS之间的潜在联系为出发点,同时基于课题组现有天然活性物质进行药物筛选,经文献调研发现青蒿素有抗肥胖效果,继而通过网络药理学方法分析预测青蒿素可能用于治疗PCOS的潜在作用靶点,并探讨其可能用于临床治疗PCOS的可行性。
本研究根据PPI网络拓扑属性分析筛选出核心靶点AKT1、MMP9、ESR1、PPARγ、MMP2等,推测这些可能是青蒿素用于治疗PCOS的潜在作用靶点。有研究表明[29],AKT1在颗粒细胞增殖中起关键作用,而其表达量的高低主要与机体雄激素水平异常有关,这会导致PCOS患者卵巢颗粒细胞正常功能受损。此外,AKT1还具有组织特异性,Song等[30]通过小鼠实验发现脂肪组织中AKT1的选择性抑制可以刺激白色脂肪组织发生褐变,从而可增加机体能量消耗发挥抗肥胖的效果。基质金属蛋白酶(matrix metalloproteinase,MMPs)是一种锌依赖性酶,可由卵巢产生,在卵泡发育和PCOS的发病机制中起重要作用[31];研究发现PCOS女性患者MMPs活性增加,其血液、卵泡液和颗粒细胞中MMP9、MMP2水平升高,高水平的MMPs会通过改变细胞外基质重塑,引起异常卵泡闭锁和卵巢基质组织增加,从而对患者的排卵和生育能力产生不良影响[32]。Barbara等[33]在对正常女性和肥胖女性血清样本中的MMP浓度对比发现,体质量增加可影响女性血清中的MMP浓度。ESR是维持卵巢颗粒细胞分化、卵泡和卵母细胞生长发育以及排卵功能的关键受体[34];ESR1是一种核激素受体,作为转录因子的激活剂发挥作用[35];Schomberg等[36]在ESR基因敲除的小鼠模型中发现ESR基因缺失会导致卵泡发育受阻,以致卵泡闭锁及无排卵现象发生。Artimani等[37]在评估PCOS患者颗粒细胞中ESR基因表达时发现,ESR mRNA的表达显著低于排卵功能正常女性,认为ESR基因的显著减少可作为颗粒细胞成熟缺陷或卵泡发育停滞的指标。ESR1也是一种与线粒体功能相关的基因,研究发现其在肥胖女性脂肪组织中有减少,Zhou等[38]在人类和啮齿动物实验中证实,脂肪组织中ESR1的表达与脂肪量呈负相关,同时ESR-α作用的降低还会损害线粒体功能,促进肥胖增加,破坏机体代谢稳态。PPARγ是一种调节脂肪细胞发育和葡萄糖稳态的核受体,主要在脂肪组织中表达[39];此外在发育阶段的卵巢颗粒细胞中表达,并可受黄体生成素(luteinizing hormone,LH)水平的影响来调节机体雌激素分泌和卵巢功能[40]。此外,Lee等[41]在PCOS患者颗粒细胞中发现PPARγ mRNA表达水平下调。胡卫红等[42]研究发现PPARγ 在PCOS患者的卵巢颗粒细胞的表达异常可能与PCOS的高雄激素血症有关。
GO生物学过程富集分析表明,青蒿素治疗PCOS的生物学功能可能与细胞增殖调控、蛋白质磷酸化和RNA聚合酶Ⅱ启动子转录的正调控等生物过程有关。KEGG通路富集分析表明,青蒿素可能通过作用于PI3K/Akt、MAPK、Ras、癌症等信号通路发挥治疗作用。有研究表明,PI3K/Akt信号通路参与调节细胞增殖分化和迁移,在卵泡发育过程中对卵巢颗粒细胞的生长和凋亡起着关键作用[43],在PCOS患者颗粒细胞中与氧化应激相关的凋亡多伴随PI3K/Akt信号下调[44]。此外,研究表明PI3K/Akt信号通路还可以调节脂肪细胞的脂解与分化,从而参与机体脂质代谢[45]。MAPK信号通路参与调节多种细胞过程,如增殖、分化、转录调控等,且该通路与卵巢颗粒细胞类固醇激素的合成有关[46]。研究发现,在PCOS女性中,异常的MAPK信号传导可导致代谢信号缺陷和卵巢雄激素分泌异常增多[47]。
为进一步探索青蒿素在PCOS治疗中的潜在分子机制,本研究将天然产物青蒿素和5个与PCOS密切相关的核心靶蛋白进行分子对接验证,寻找二者之间存在的最佳结合位点以及评估它们之间的结合能力。验证结果显示,青蒿素与核心靶蛋白之间能够较好结合。
综上所述,本研究采用网络药理学方法分析天然物青蒿素用于治疗PCOS的潜在作用靶点,其机制可能主要涉及PI3K/Akt、MAPK、内分泌抵抗等信号通路。这些信息为后续青蒿素用于治疗PCOS的实验验证提供了重要理论依据。
-
图 2 三种铁死亡诱导剂分别联用hcGEM对胰腺癌PANC-1细胞存活率的影响
A.hcGEM-Era联合用药组;B.hcGEM-SASP联合用药组;C.hcGEM-Art联合用药组;*P<0.05, 与单用hcGEM组比较
图 3 三种铁死亡诱导剂分别联用hcGEM对胰腺癌PANC-1细胞的效应-联合指数曲线图
A.hcGEM-Era联用组;B. hcGEM-SASP联用组;C.hcGEM-Art联用组
表 1 hcGEM联合三种铁死亡诱导剂对胰腺癌PANC-1细胞作用时的IC50值
组别 摩尔浓度比 IC50(μmol/L,hcGEM) hcGEM-Era联合组 1∶0.25 0.140 3±0.009 1 1∶1 0.340 2±0.018 3 1∶4 0.091 3±0.005 1 1∶16 0.083 3±0.002 5 hcGEM-SASP联合组 1∶0.25 0.297 5±0.016 1 1∶1 0.240 2±0.021 2 1∶20 0.120 8±0.008 9 1∶400 0.092 6±0.006 7 hcGEM-Art联合组 1∶0.5 0.366 4±0.018 7 1∶1 0.344 4±0.026 3 1∶4 0.249 3±0.015 7 1∶16 0.154 6±0.013 5  下载: 导出CSV
下载: 导出CSV
-
[1] BRAY F, FERLAY J, SOERJOMATARAM I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA A Cancer J Clin,2018,68(6):394-424. doi: 10.3322/caac.21492 [2] MIZRAHI J D, SURANA R, VALLE J W, et al. Pancreatic cancer[J]. Lancet,2020,395(10242):2008-2020. doi: 10.1016/S0140-6736(20)30974-0 [3] PAROHA S, VERMA J, DUBEY R D, et al. Recent advances and prospects in gemcitabine drug delivery systems[J]. Int J Pharm,2021,592:120043. doi: 10.1016/j.ijpharm.2020.120043 [4] General Office of National Health Commission;. 胰腺癌诊疗指南(2022年版)[J]. 临床肝胆病杂志, 2022, 38(5):1006-1030. doi: 10.3969/j.issn.1001-5256.2022.05.007 [5] LEI G, ZHUANG L, GAN B Y. Targeting ferroptosis as a vulnerability in cancer[J]. Nat Rev Cancer,2022,22(7):381-396. doi: 10.1038/s41568-022-00459-0 [6] CHEN M J, LI X N, DU B L, et al. Upstream stimulatory factor 2 inhibits erastin-induced ferroptosis in pancreatic cancer through transcriptional regulation of pyruvate kinase M2[J]. Biochem Pharmacol,2022,205:115255. doi: 10.1016/j.bcp.2022.115255 [7] LO M, LING V, LOW C, et al. Potential use of the anti-inflammatory drug, sulfasalazine, for targeted therapy of pancreatic cancer[J]. Curr Oncol,2010,17(3):9-16. doi: 10.3747/co.v17i3.485 [8] WANG K, ZHANG Z Y, WANG M, et al. Role of GRP78 inhibiting artesunate-induced ferroptosis in KRAS mutant pancreatic cancer cells[J]. Drug Des Devel Ther,2019,13:2135-2144. doi: 10.2147/DDDT.S199459 [9] CHEN X, KANG R, KROEMER G, et al. Targeting ferroptosis in pancreatic cancer: a double-edged sword[J]. Trends Cancer,2021,7(10):891-901. doi: 10.1016/j.trecan.2021.04.005 [10] WOOD L D, CANTO M I, JAFFEE E M, et al. Pancreatic cancer: pathogenesis, screening, diagnosis, and treatment[J]. Gastroenterology,2022,163(2):386-402.e1. doi: 10.1053/j.gastro.2022.03.056 [11] HAN H, LI S, ZHONG Y, et al. Emerging pro-drug and nano-drug strategies for gemcitabine-based cancer therapy[J]. Asian J Pharm Sci,2022,17(1):35-52. doi: 10.1016/j.ajps.2021.06.001 [12] LI C, YIN X Z, LIU Z J, et al. Emerging potential mechanism and therapeutic target of ferroptosis in PDAC: a promising future[J]. Int J Mol Sci,2022,23(23):15031. doi: 10.3390/ijms232315031 [13] STOCKWELL B R. Ferroptosis turns 10: emerging mechanisms, physiological functions, and therapeutic applications[J]. Cell,2022,185(14):2401-2421. doi: 10.1016/j.cell.2022.06.003 [14] CHEN X, KANG R, KROEMER G, et al. Broadening horizons: the role of ferroptosis in cancer[J]. Nature reviews Clinical oncology,2021,18(5):280-296. doi: 10.1038/s41571-020-00462-0 [15] DAHER B, PARKS S K, DURIVAULT J, et al. Genetic ablation of the cystine transporter xCT in PDAC cells inhibits mTORC1, growth, survival, and tumor formation via nutrient and oxidative stresses[J]. Cancer Res,2019,79(15):3877-3890. doi: 10.1158/0008-5472.CAN-18-3855 [16] OHMAN K A, HASHIM Y M, VANGVERAVONG S, et al. Conjugation to the sigma-2 ligand SV119 overcomes uptake blockade and converts dm-Erastin into a potent pancreatic cancer therapeutic[J]. Oncotarget,2016,7(23):33529-33541. doi: 10.18632/oncotarget.9551 [17] WANG K, ZHANG Z Y, TSAI H I, et al. Branched-chain amino acid aminotransferase 2 regulates ferroptotic cell death in cancer cells[J]. Cell Death Differ,2021,28(4):1222-1236. doi: 10.1038/s41418-020-00644-4 [18] ELING N, REUTER L, HAZIN J, et al. Identification of artesunate as a specific activator of ferroptosis in pancreatic cancer cells[J]. Oncoscience,2015,2(5):517-532. doi: 10.18632/oncoscience.160 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3972
- HTML全文浏览量: 1795
- PDF下载量: 23
- 被引次数: 0

