

 下载:
下载:

-
药物在体内的跨质膜运输,大部分由药物转运体介导。这些药物转运体一般分为溶质载体(SLC)家族和ATP结合盒(ABC)家族。这些转运体不仅介导治疗药物的跨膜转运,还转运各种内源性化合物。因此,除了参与药物的处理,使其产生临床疗效或毒性外[1],这些转运蛋白还通过调节内源性化合物的转运在维持细胞稳态方面发挥重要作用。转运体是体内位于细胞膜上的功能性膜蛋白,是介导物质和能量跨膜转运的关键结构,在药物的吸收、分布、代谢、排泄动力学过程中发挥重要作用。近年来,许多研究表明,多数药物性损伤可能与机体某部位上转运蛋白的表达改变有关。转运体是体内位于细胞膜上的功能性膜蛋白,是介导物质和能量跨膜转运的关键结构。转运体的异常表达或功能缺失是引起药物毒性的重要原因之一。
药物的吸收主要在肠道中进行,其中,肠道转运体发挥着重要作用。近年来研究发现,肠道菌群可以通过影响肠道转运体蛋白的表达来影响吸收转运。同时,中药成分也可以通过调控肠道菌群来影响肠道转运体。明确中药和肠道菌群对转运体表达的影响,有利于提高药物的安全性和有效性,指导临床合理用药。
在我国,中医药已有几千年的应用历史,在改善人体亚健康、防治重大疾病等方面发挥重要作用。有研究表明,中药(天然药物提取物和中草药配方)可以显著改善肠道菌群的组成和代谢功能,有助于维持肠道菌群稳态,从而调节物质代谢与吸收转运。例如,冬凌草素[2]被证明可以通过改变肠道微生物群和促进肝尿素循环来减少肝损伤;化瘀排痰方、葛根芩连汤可显著上调Claudin-1、occludin和ZO-1的表达,修复肠黏膜屏障,防止慢性炎症的发生[3]。
人体肠道中的肠道菌群是肠道微生态系统的重要组成部分。研究发现胃肠道中至少包含9个细菌门,其中,双歧杆菌属、乳酸杆菌属、拟杆菌属、肠杆菌属、肠球菌属、梭菌属等为胃肠道主要的优势菌属。正常情况下[4],一个组合合理、协调平衡的胃肠道菌群生态体系具有重要的生理作用,包括生物屏障作用、营养代谢作用、免疫作用、抗衰老及抗肿瘤作用等。因此,人类肠道微生物群已经成为控制肠道屏障和新陈代谢的关键 [5]。目前已有一些研究对肠道菌群如何调控药物代谢进行了探索,但其具体机制仍不清楚。本综述旨在介绍中药使用过程中,中药成分如何影响肠道菌群,以及肠道菌群对转运蛋白的影响,揭示肠道菌群直接或间接发挥作用影响转运体的表达。
-
肠道菌群已被证实会影响多种口服药物的药动学过程,其除了直接影响药物的肠道代谢转化外,还可以通过调节转运体的表达和活性,间接影响药物作用。
-
紧密连接蛋白是上皮细胞和内皮细胞中细胞间连接复合体的最顶端成分。它们分离顶端和基底外侧细胞表面域(栅栏功能),并抑制溶质和水通过细胞旁空间流动(屏障功能)[6]。其由整合膜蛋白claudins、occludin和连接黏附分子(JAM)以及许多外周膜蛋白形成,包括膜相关鸟苷酸激酶(MAGUK)家族、ZO-1、ZO-2、ZO-3、具有反向取向的膜相关鸟糖苷酸激酶1(MAGI-1)、细胞极性分子ASIP/PAR-3、PAR-6和非典型蛋白激酶C(PKC)[7]。Occludin是首个被报道的紧密连接的整合膜蛋白,并且在最顶端的基底外侧膜上表达最普遍。此外,occludin还直接参与紧密连接屏障和栅栏功能[8]。
肠道菌群与肠黏膜机械屏障中紧密连接蛋白的表达关系密切。肠道菌群失调后,会引起肠黏膜紧密连接蛋白Claudin-1、occludin和ZO-1的表达降低,紧密连接被破坏,发生肠黏膜细胞核易位等现象[9]。研究发现,剧烈运动可引起胃肠痉挛、腹泻等肠道症状,这可能是由于运动引起体温升高导致肠道热损伤和菌群变化[10],使得肠道中有害菌直接与肠上皮细胞表面分子结合,改变紧密连接蛋白occludin、ZO-1等的表达,破坏紧密连接结构,损害细胞骨架,从而增大细胞间“空隙”,增加细胞旁通透性,损伤肠黏膜屏障,引发内毒素血症。瘤胃球菌已被证明可以促使杯状细胞分泌黏蛋白,阻止有害细菌的入侵[11]。类副杆菌作为有益微生物,可以降低促炎细胞因子的水平,形成稳态定植以维持肠黏膜屏障[12]。这些益生菌[13]是维持肠道细菌之间生物对抗性的主要菌种,有助于肠道兼性细菌的生长,增强细胞免疫和体液免疫,减少肠上皮细胞的炎症反应,增强肠道防御功能,并在调节肠道菌群组成方面发挥重要作用。
-
外排转运蛋白,如P-糖蛋白、ABCB1或MDR1,可以主动将广泛的化学性化合物泵出细胞,保护肠黏膜免受异生物质和代谢产物(包括药物)的影响。缺乏P-糖蛋白,可导致异生物质在细胞的积累和毒性增加,并伴有严重的副作用。肠道细菌可以调节宿主P-糖蛋白的功能[14]。例如,阿克曼菌可以参与肠道炎症的调节[15, 16],通过上调短链脂肪酸(SCFA)的产生使得小鼠肠道中P-gp的表达下降。革兰阴性肠杆菌和克雷伯菌可产生外膜囊泡(OMV),其作用于TL受体,改变CYP3A和P-gp的表达[17]。Caco-2单层膜和葡聚糖硫酸钠诱导结肠炎小鼠模型实验显示,嗜酸乳杆菌、鼠李糖乳杆菌通过磷酸肌苷3-激酶和ERK1/2 MAPK途径刺激P-gp活性,同时可使回肠和结肠中mdr1a/P-gp mRNA和蛋白表达增加2~3倍[18]。肠道菌群通过影响外排转运蛋白的表达来影响肠道的吸收转运功能,为之后中药成分调控肠道菌群来影响肠道吸收转运奠定基础。
-
中药通过肠道菌群发挥吸收转运的调节作用,主要包括改变肠道菌群组成、影响肠道菌群代谢以及影响肠道屏障功能3种途径。
-
临床无菌模型和粪便移植疗法证明,肠道菌群可以靶向细菌,达到中医治疗疾病的目的,主要体现在中药有效成分对肠道菌群结构、组成和代谢产物的影响。例如,白藜芦醇可以重塑不同水平肠道菌群的多样性和组成[19]。在门水平上,厚壁细菌数量显著增加,拟杆菌数量显著减少;在科水平上,丹毒科增加;在属水平上,奥氏菌含量增加。白芦醇通过丰富以上肠道菌群的结构,从而影响肠道屏障的紧密连接功能[19],同时通过下调炎症因子(IL-1、TNF- α、MyD88和TLR-4)改善肝脏炎症,进而改进吸收转运的功能。鸡腿菇多糖是食用菌Coprinus comatus的多糖提取物,已被证明具有类似益生元的作用,可以增加肠道微生物群的多样性(厚壁菌和乳酸菌科相对丰度显著较高)[20],通过丰富厚壁菌增加紧密连接蛋白和乳酸杆菌来激活P-gp活性,从而影响肠道吸收转运。此外,人参提取物和酸枣种子(GS)显著提高了大鼠乳酸菌和双歧杆菌的相对丰度,降低了链球菌、大肠杆菌-志贺杆菌、绒毛杆菌和肠球菌的相对丰度,提示GS提取物可能通过平衡肠道微生物群的结构和多样性[21]进而恢复吸收转运的功能,是一种有前景的酒精性肝病(ALD)治疗药物。
此外,一些中药[22]可以直接抑制微生物的生长。例如,肉桂精油可以抑制大肠杆菌和金黄色葡萄球菌。粪便微生物学结合16S rDNA分析表明,加味逍遥散显著改变了肠道微生物群的组成[23],增加有益菌,减少有害菌的组成,并影响了11种差异代谢产物(初级胆汁酸生物合成、苯丙氨酸代谢、泛酸和辅酶A生物合成、代谢途径、胆固醇代谢、胆汁分泌等),从而达到影响吸收转运的目的。小檗碱是从草药中提取的一种天然异喹啉生物碱,是黄连和小檗的主要活性成分。小檗碱处理后,大鼠肠道菌群的多样性和丰富度发生了变化[24],拟杆菌的丰度提高,乳酸菌显著上调,变形杆菌和疣状微生物的丰度降低。小檗碱通过重塑肠道菌群结构来改变紧密连接蛋白和外排转运蛋白的表达,进而影响肠道吸收转运功能。在没食子酸(GA)对多重耐药大肠杆菌的杀菌活性研究中表明,GA具有杀菌活性,并能抑制细菌生物膜的形成。进一步的形态学改变和外排泵基因表达分析证实[25],GA损伤了大肠杆菌的外膜和内膜,并抑制了与膜通透性有关的acrA、acrB、tolC、acrD和acrF的mRNA表达。Yang等[26]发现,芦丁可通过增加肠道菌中Bacillus 、Bacteroides 和Veillonella 的丰度,促进β-葡萄糖苷酶活性,使其转化成易吸收的代谢物。
肠道内产生的某些酶会使肠上皮受到损伤。白藜芦醇、菊粉、大黄、槲皮素等的中药制剂,如四脉丸、金芪降糖T片、黄连解毒司等均可增加阿克曼菌的丰度,降低大肠杆菌的丰度。因为大肠杆菌不利于维持肠道屏障的完整性,其产生的代谢酶StcE会分解黏蛋白[27],增加肠道通透性,诱发肠道炎症。上述的中药成分可以降低大肠杆菌的丰度,从而维护人体的肠道屏障,进而改善吸收转运功能。Shen等[28]通过粪便温孵人参皂苷Rb1体外代谢研究和ABXco-2细胞单层转运研究,证明人参多糖(GP)减轻了葡聚糖硫酸钠(DSS)引起的结肠炎样症状,并通过增强微生物的糖基化、肠上皮细胞对Rb1的吸收来提高Rb1的全身暴露量。这些发现进一步证明了多糖与肠道菌群相互作用在中药传统汤剂整体效用中的重要作用。Guo等[29]比较了对照组和伪无菌大鼠组给药三七皂苷后的代谢产物数量,发现对照组有73种代谢产物,伪无菌大鼠中仅检测到11种代谢产物,表明肠道菌群参与体内三七皂苷的生物转化。Song等[30]发现大黄提取物和大黄酸均可在肠道菌群介导下发生还原、水解或进一步乙酰化等生物转化。Huang等[31]也证明肠道菌群介导的大黄中蒽醌和蒽酮还原、水解、乙酰化、氧化、去甲基化、甲基化、羟基化、脱羟基化等反应在其体内生物活性中起重要作用。
-
相关研究证明,肠道菌群[32]可以通过水解、氧化、还原和异构化反应产生低极性和相对稳定的中药代谢物,从而加速肠道吸收,提高中药的生物利用度。例如,Racova等[33]采用体外粪便培养方式研究黄酮类化合物的代谢产物,发现其主要代谢途径为水解、去糖基化、羟基化等,随后粪便细菌形成的代谢物被肠道酶进一步发生谷胱甘肽偶联或葡萄糖醛酸化,表明肠道菌与黄酮类化合物的代谢存在联系。有研究表明,肠道微生物代谢产物胆汁酸(BA)能通过抑制P-gp ATP酶来调节P-gp活性[34],Telbisz等[35]证明糖胆酸、牛磺胆酸和胆酸等多种BA可显著降低基础BCRP ATP酶活性;短链脂肪酸丁酸盐促进上皮屏障功能,通过诱导编码紧密连接(TJ)成分的基因来巩固肠道屏障[36],从而影响肠道吸收转运。当肠道微生物群发生紊乱时,肠道微生物群代谢物(短链脂肪酸、三甲胺N-氧化物、BA、脂多糖等)[37]会相应地发生变化,肠道屏障会被破坏。传统中药可以通过干预肠道微生物群代谢物来改善肠道环境,从而影响肠道吸收转运。例如,生物碱(尤其是小檗碱)被证明可以调节胆固醇到BA的转化并促进粪便BA排泄[38]。脾胆健清汤的主要成分为黄芪、黄连、黄芩、苍术、丹参、荔枝。脾胆健清汤对糖尿病的临床治疗有良好的干预作用,对于吸收转运功能有极大的影响。其作用机制为,在属水平上,脾胆健清汤提高了乳酸菌、布鲁菌、拟杆菌、脱硫弧菌和阿克曼菌的相对丰度,降低了普雷沃菌的相对丰度,从而增加短链脂肪酸的生成[39]。相关分析表明[40],脾胆健清汤对色氨酸代谢、组氨酸代谢和三羧酸(TCA)循环途径的调节作用与乳杆菌、拟杆菌和阿克曼细菌的丰度变化有关。因此,脾胆健清汤通过对肠道菌群代谢物的影响治疗糖尿病,同时影响吸收转运功能。
-
肠黏膜屏障是存在于肠道中的一种高选择性的功能屏障系统,可以防止有害物质,如病原微生物和内毒素通过肠黏膜,同时选择性地吸收肠内营养物质。也在免疫防御和维持肠黏膜完整性方面发挥重要作用[41]。肠黏膜损伤、炎症因子升高与代谢性疾病(糖尿病、酒精性脂肪肝等)有关。中药通过增加肠道黏液和紧密连接蛋白来减轻代谢性炎症[42],进而改善吸收转运功能。许多研究表明,中药可以通过改善肠黏膜微循环、修复肠黏膜完整性、降低肠黏膜通透性来维持机械屏障功能。一项研究表明[43],黄芩苷可提高粪便中丁酸盐的水平,丁酸盐水平的增加可提高UC大鼠ZO-1、Occludin和Muc2的表达,调节Th17/Treg平衡,减少肠黏膜损伤,增强肠道屏障功能,从而影响肠道吸收。姜黄多糖(TPS)是从中国植物药姜黄中提取的,在用TPS处理的UC小鼠中,盲肠色氨酸分解代谢产物吲哚-3-乙酸(IAA)及其配体芳烃受体(AhR)的表达显著增加。据推测,TPS通过激活AhR上调上皮紧密连接蛋白肠屏障功能发挥保护作用,从而影响肠道吸收功能[44]。单宁酸和黄连素是从中草药(大黄和黄连)中提取的两种活性成分,可以恢复紧密连接相关蛋白的表达,对UC发挥治疗作用,增强肠道屏障功能,对肠道吸收产生影响[45]。白头翁汤(PD)是临床上常用于治疗UC的中药配方,PD能缓解UC小鼠的症状,PD高剂量治疗组(PDHT)效果最佳。PDHT组中[46]Occludin、Occluden小带-1 (ZO-1)、Claudin1、Claudin5、G蛋白偶联受体43 (GPR43)蛋白的表达水平以及ZO-1和Occludin mRNA的相对表达量均显著升高,从而增强肠道屏障的功能,对肠道吸收转运产生影响。
-
近年来,随着肠道微生物在健康调控和代谢控制中的作用逐步得到确认,肠道微生物相关健康科学得到了飞速发展。利用中药优化肠道菌群结构进而提升人类健康水平以及探究肠细胞上机械屏障和外排转运蛋白对肠道吸收转运机制的影响具有重要意义。肠道菌群对转运蛋白的影响呈现双面性,了解肠道菌群对转运蛋白的影响可能有助于开发毒性较小的转运蛋白调节剂,指导临床合理用药。例如,Ghosh等[47]研究肠道微生物代谢物尿石素对逆转5-氟尿嘧啶耐药结肠癌的影响和机制时发现,尿石素及其结构类似物能下调MDR5、BCRP、MRP1和MRP2等药物转运蛋白的表达和活性,使结肠癌细胞化学敏化,5-氟尿嘧啶的外排减少,有利于癌症治疗。
然而,尚有一些问题值得进一步探讨。首先,中药优化肠道菌群结构、调节肠道机械屏障和外排转运蛋白是否能提高合理用药的效率;其次,具体调节机制仍需要深入挖掘;最后,除了转运体表达外,是否有其他肠道内成分可以起到影响药物排出的作用。另外,肠道菌群除了受中药成分影响外,与日常生活方式、运动习惯、饮食结构也有很大关系,需要进一步的研究。
Research progress on the regulation of intestinal flora and effect on intestinal absorption and transport by TCM components
-
摘要: 概括介绍中药成分调控肠道菌群影响肠道吸收转运作用的国内外研究进展,为后续以肠道菌群为靶点研究临床合理用药提供帮助。通过查阅近年来肠道菌群与肠道吸收转运方面的文献,分析总结现有相关研究,阐述中药成分调控肠道菌群对药物吸收转运的作用机制。研究发现中药成分改变肠道菌群进而影响肠道屏障功能和吸收转运,对临床合理用药具有重要意义。Abstract: The domestic and international research progress on the regulation of gut microbiota by Traditional Chinese Medicine (TCM) ingredients and their impact on intestinal absorption and transportation were summarized, which provided assistance for subsequent clinical rational drug use targeting gut microbiota. Literature on the relationship between gut microbiota and intestinal absorption and transportation in recent years were reviewed and analyzed, and the mechanism of TCM ingredients regulating gut microbiota on drug absorption and transportation was elucidated. Research has found that TCM ingredients alter gut microbiota, thereby affecting intestinal barrier function and absorption of transport proteins, which is of great significance for rational clinical medication.
-
艰难梭菌的感染率近年来有所提高[1],其孢子很难根除,并且很容易通过医护人员传播,是医院、养老院和一些健康护理机构出现腹泻感染的原因之一[2]。目前针对艰难梭菌的可用治疗方法都有一些缺点,例如成本过高、耐药性的发展和选择性问题。 虽然轻度至中度的艰难梭菌病例目前可以通过可用的治疗方法进行控制,但对严重的病例可能需要使用单一常规药物或联合疗法进行长期抑制治疗,如果不成功,则要采用非常规和(或)侵入性的手段,如粪便移植和结肠切除术[3]。因此,临床上对药物研发需求越来越大,寻求一种更有效的替代疗法迫在眉睫[1]。
含有喹啉的杂芳族化合物具有多种生物和药理活性,例如,抗疟疾[4]、抗结核病[5]、抗炎[6]、抗癌[7]、抗高血压[8]以及抗菌[9-11]。Musiol 等[12]曾报道 8-羟基喹啉衍生物的抗真菌活性。此外,8-羟基喹啉衍生物显示出对神经退行性疾病如阿尔茨海默病良好活性[13]。最近,研究发现8-羟基喹啉衍生物具有抗金黄色葡萄球菌的潜在作用[14]。我们的小组最近通过药物数据库筛选发现,溴氧喹啉(5,7-二溴喹啉-8-酚)对艰难梭菌具有强效抗菌活性。基于此有前景的化合物,我们进行了一系列8-羟基喹啉类似物的设计、合成和抗艰难梭菌活性研究。
1. 目标化合物的设计
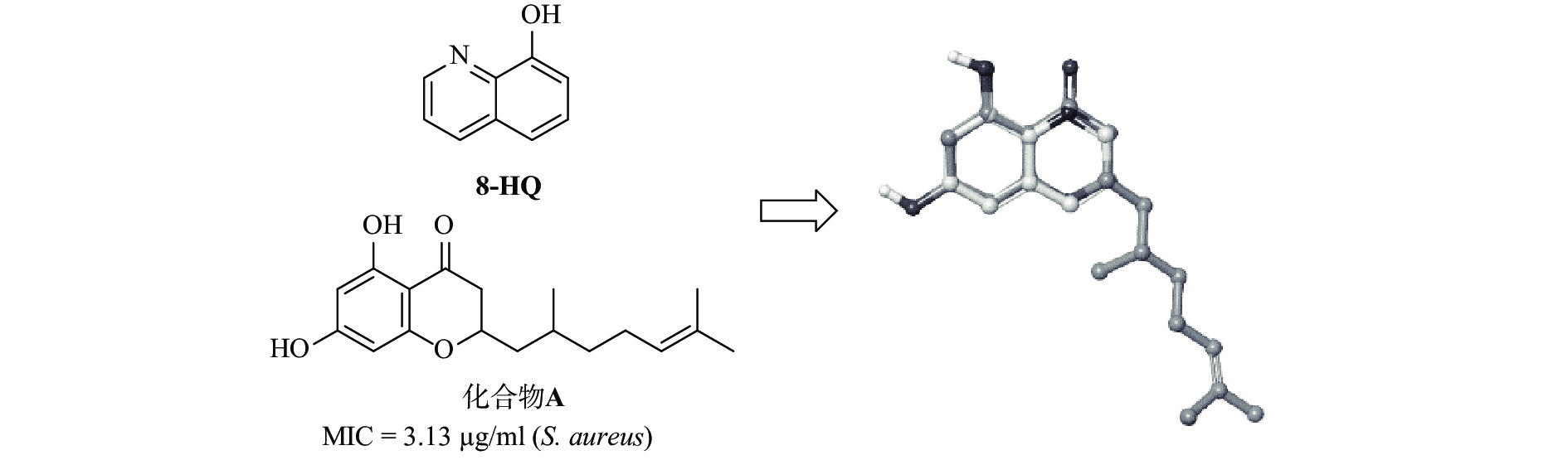
早期研究发现,黄酮化合物 A(图1)具有良好的抗菌活性,对金黄色葡萄球菌的最小抑菌浓度(MIC)为 3.13 μg/ml [15],而从8-羟基喹啉与化合物A的叠合图(图1)可见,8-羟基喹啉骨架具有与黄酮类化合物相似的药效基团。为了探索取代基的功能并研究详细的 SAR,我们设计并合成了一系列在8-羟基喹啉5、 6和7 位具有芳香取代基的化合物。
2. 材料和方法
2.1 试剂和仪器
化学原料均为市售分析纯;Bruker AM-400 (400 MHz) 核磁共振仪,TMS 作为内标,化学位移与偶合常数分别用 ppm 和 Hz 表示 ;Agilent 6538 UHD Accurate-Mass Q-TOF LC/MS高分辨质谱(HRMS)仪。
2.2 6-位取代目标化合物的合成
4-溴-2-甲氧基苯胺通过 Skraup 合成[16] ,在 H2SO4、3-硝基苯磺酸钠和甘油中回流,得到中间体 1(见图2)。随后,甲基保护基被脱除,再在DIPEA存在下用MOMCl保护,得到化合物 2。此外,化合物 1 还经 Suzuki 反应与芳基硼酸偶联反应得到中间体 3a-j 。 化合物2与苯乙烯偶联,得到化合物 4a,然后使用 1 mol/L 的盐酸脱保护,得到化合物 5a。其中,化合物 3a-g 和 3i-j使用 HBr (48%) 在回流下脱保护,以较高的收率得到化合物 6a-g 和 6i-j,化合物 3h 用 BBr3 处理得到化合物 6h。
 图 2 化合物1~6的合成注:a. H2SO4, H2O, 3-硝基苯磺酸钠, 甘油, 加热回流, 6 h; b. HBr, 回流, 8 h; c. 苯乙烯, Pd(dppf)Cl2, 碳酸钾, 二氧六环, 100 ℃, 过夜; d. 取代的芳基硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 95 ℃, 2 h; e. 三溴化硼, 二氯甲烷, 0 ℃到室温, 过夜; f. MOMCl, DIPEA, 二氯甲烷, 室温, 过夜; g. 盐酸水溶液, THF, i-PrOH, 室温过夜
图 2 化合物1~6的合成注:a. H2SO4, H2O, 3-硝基苯磺酸钠, 甘油, 加热回流, 6 h; b. HBr, 回流, 8 h; c. 苯乙烯, Pd(dppf)Cl2, 碳酸钾, 二氧六环, 100 ℃, 过夜; d. 取代的芳基硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 95 ℃, 2 h; e. 三溴化硼, 二氯甲烷, 0 ℃到室温, 过夜; f. MOMCl, DIPEA, 二氯甲烷, 室温, 过夜; g. 盐酸水溶液, THF, i-PrOH, 室温过夜为了进一步研究8-羟基喹啉衍生物的构效关系,我们还设计并合成了)一系列6位杂环取代的化合物, 如图3 所示。首先,中间体1或相应的硼酸与市售芳基硼酸或卤代物通过Suzuki反应偶联得到7a-i,然后在氢溴酸(48%)中回流脱甲基以高收率得到化合物8a-i。
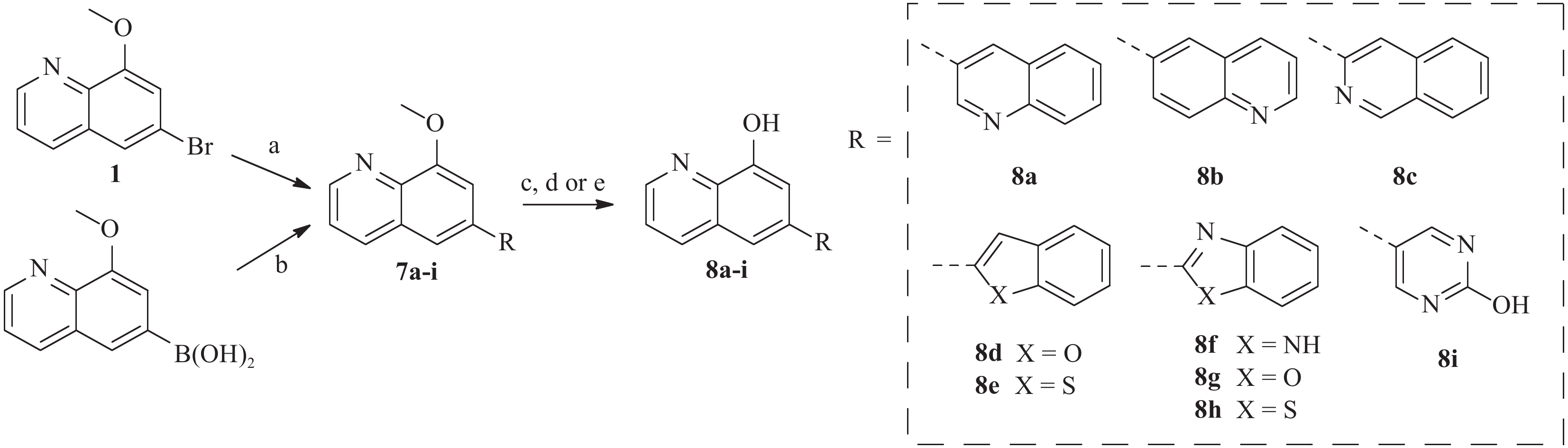 图 3 化合物8的合成注:a. 取代的芳基硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 100 ℃, 过夜; b. 芳基溴代物, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 100 ℃, 过夜; c. 氢溴酸, 回流, 过夜; d. 三溴化硼, 二氯甲烷, 0 ℃到室温, 过夜; e. 盐酸水溶液, 四氢呋喃, 室温, 过夜
图 3 化合物8的合成注:a. 取代的芳基硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 100 ℃, 过夜; b. 芳基溴代物, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 100 ℃, 过夜; c. 氢溴酸, 回流, 过夜; d. 三溴化硼, 二氯甲烷, 0 ℃到室温, 过夜; e. 盐酸水溶液, 四氢呋喃, 室温, 过夜2.3 5-位和7-位取代目标化合物的合成
为了研究5和7位的构效关系,我们还设计合成了一系列在5和7位具有不同取代基的化合物。 如图4 所示,市售的 7-溴-8-羟基喹啉(9)经MOM和甲基保护,分别得到中间体10a和10b。 随后,这些中间体与苯乙烯和苯硼酸偶联,分别得到中间体11a 和11b。 最后,化合物 17a 和 17b 分别经 1 mol/L盐酸水溶液和氢溴酸水溶液(48%)脱保护,得到所需的 8-羟基喹啉类似物18a 和18b。 此外, 5 位取代化合物的合成如图5 所示。首先,将市售的8-羟基喹啉在碳酸钾存在下与碘甲烷反应,随后在甲醇中与溴素在室温下反应得到中间体12[17]。随后,化合物12按照与前述相同的方法合成13。
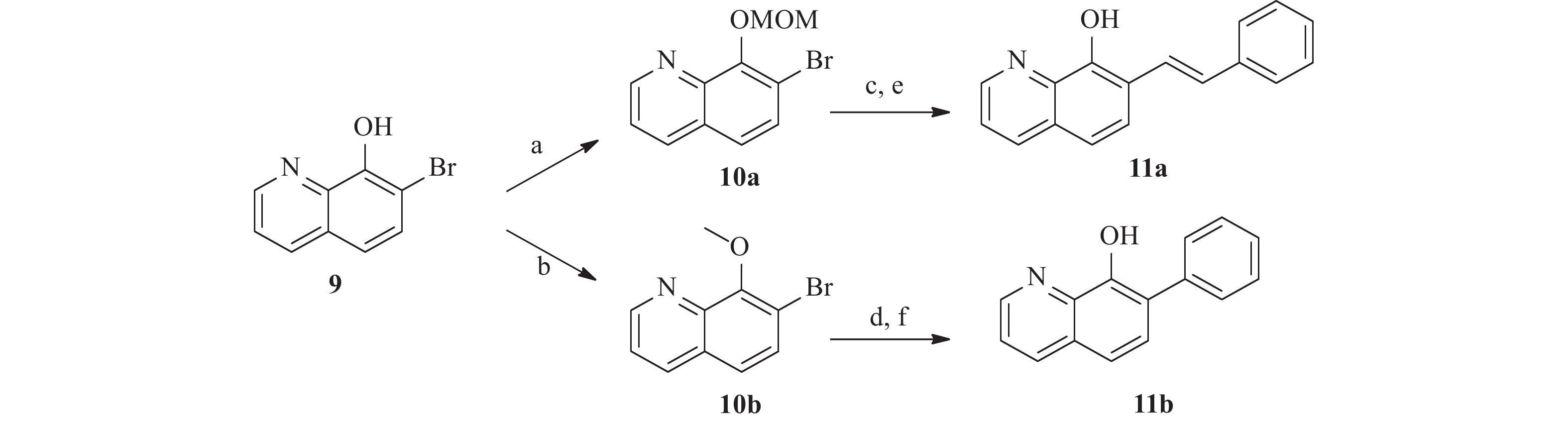 图 4 化合物11的合成注:a. MOMCl, DIPEA, 二氯甲烷, 0 ℃室温搅拌过夜; b. 碘甲烷, 碳酸钾, 0 ℃室温搅拌, 2 h; c. 苯乙烯, Pd(dppf)Cl2, 碳酸钠, DMF, 100 ℃, 过夜; d. 苯硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 95 ℃, 过夜; e. 盐酸水溶液, 四氢呋喃, 室温搅拌过夜; f. 氢溴酸, 回流, 8 h
图 4 化合物11的合成注:a. MOMCl, DIPEA, 二氯甲烷, 0 ℃室温搅拌过夜; b. 碘甲烷, 碳酸钾, 0 ℃室温搅拌, 2 h; c. 苯乙烯, Pd(dppf)Cl2, 碳酸钠, DMF, 100 ℃, 过夜; d. 苯硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 95 ℃, 过夜; e. 盐酸水溶液, 四氢呋喃, 室温搅拌过夜; f. 氢溴酸, 回流, 8 h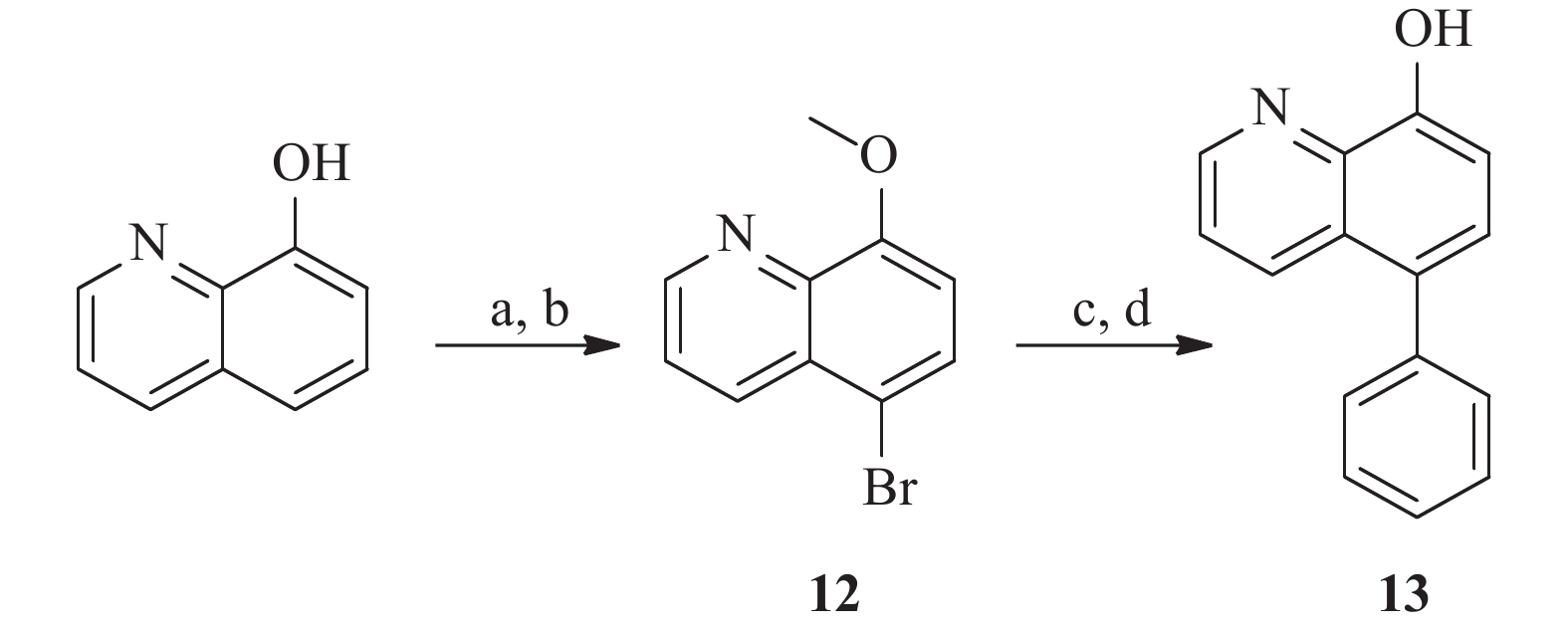 图 5 化合物13的合成注:a. 碘甲烷, 碳酸钾, 0 ℃至室温, 4 h; b. 溴素, 甲醇, 室温搅拌6 h; c. 氢溴酸, 回流, 10 h; d. 苯硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 95 ℃, 过夜
图 5 化合物13的合成注:a. 碘甲烷, 碳酸钾, 0 ℃至室温, 4 h; b. 溴素, 甲醇, 室温搅拌6 h; c. 氢溴酸, 回流, 10 h; d. 苯硼酸, Pd(PPh3)4, 碳酸钾, 水, 二氧六环, 95 ℃, 过夜3. 结果
3.1 8-羟基喹啉衍生物的抗艰难梭菌活性评价
所有目标化合物及部分中间体均针对两种菌株 BAA1875 和 R20291 的艰难梭菌进行了活性测试。基于生物活性(见表1),SAR总结如下:①8位的游离羟基对艰难梭菌的抑制活性起着非常重要的作用,其甲基化会大幅降低活性(化合物1和3c分别对BAA-1875为大于256 μg/ml)。② 6 位的取代对于活性很重要,体积大的基团(例如萘基)对活性有利(萘基 6f > 苯基 6a )。此外,对于 6 位萘取代的化合物,2-萘基优于1-萘基(6f vs 6h),且带有 OH 或 OCH3 取代基的萘环也会降低活性(5b-c, 6i-j)。③对于6位杂环取代的类似物,引入杂原子后会导致活性整体下降,且六六并环( 8a-c )的活性显著好于五六并环( 8d-h )。 ④对于7位的替换,大基团对活性不利。溴原子是最佳取代,随着取代基大小的增加,活性略有下降(9>11b>11a)。 ⑤5 位苯环取代依旧可以保持活性(13)。
表 1 针对两种菌株BAA1875和R20291的抗艰难梭菌活性 (MIC, µg/ml)化合物 BAA 1875a R20291a 化合物 BAA 1875a R20291a 1 >256 >256 8a 4 2 3c >256 >256 8b 8 8 5a 4 2 8c 8 4 5b 32 32 8d >256 >256 5c 8 8 8e >256 >256 6a 1 4 8f 32 32 6b 1 4 8g 64 32 6c >256 >256 8h 64 16 6d >256 >256 8i 32 32 6e 8 8 9 0.5 0.25 6f 0.5 0.25 11a 4 4 6g 2 2 11b 2 1 6h 8 4 13 0.5 0.25 6i 16 8 万古霉素 0.25 0.125 6j 16 16 注:a.化合物起始浓度256 µg/ml 3.2 部分8-羟基喹啉衍生物的MIC和MBC测试结果
进一步测试了部分8-羟基喹啉衍生物针对艰难梭菌R20291的最小杀菌浓度(MBC)(表2),所测试化合物的最小杀菌浓度(MBC)和MIC相当,表明这些化合物能够有效地杀死艰难梭菌,具有更强的抑菌作用。在这些化合物中,化合物 6f 表现出良好的活性,这使其成为可进一步开发的良好先导化合物。
表 2 部分8-羟基喹啉衍生物的MIC和MBC测试结果(µg/ml)化合物 R20291 MIC MBC 5a 3.2 3.2 6e 6.4 6.4 6f 0.1 0.2 6h 3.2 3.2 9 0.1 0.2 11a 3.2 3.2 13 0.4 0.4 4. 讨论
基于重要的抗艰难梭菌骨架8-羟基喹啉,我们设计并合成了一系列8-羟基喹啉衍生物,所有目标化合物均经过了核磁和质谱确证结构,纯度均高于95%。 针对艰难梭菌菌株BAA1875和R20291的抑菌试验显示,多个化合物具有强效的抑菌作用,且MIC 与MBC 非常接近,表明这些化合物还具有强效的杀菌作用。这使它们成为可进一步开发的良好先导化合物,后续的结构优化工作尚在进行之中。
-
[1] LIU X D. Overview: role of drug transporters in drug disposition and its clinical significance[J]. Adv Exp Med Biol, 2019, 1141:1-12. [2] HONG M K, LIU H H, CHEN G H, et al. Oridonin alters hepatic urea cycle via gut microbiota and protects against ace-taminophen-induced liver injury[J]. Oxid Med Cell Longev, 2021, 2021:3259238. [3] LIU M, SHI W, HUANG Y F, et al. Intestinal flora: a new target for traditional Chinese medicine to improve lipid metabolism disorders[J]. Front Pharmacol, 2023, 14:1134430. doi: 10.3389/fphar.2023.1134430 [4] FUJIMURA K E, SLUSHER N A, CABANA M D, et al. Role of the gut microbiota in defining human health[J]. Expert Rev Anti Infect Ther, 2010, 8(4):435-454. doi: 10.1586/eri.10.14 [5] DE VOS W M, TILG H, VAN HUL M, et al. Gut microbiome and health: mechanistic insights[J]. Gut, 2022, 71(5):1020-1032. doi: 10.1136/gutjnl-2021-326789 [6] GUMBINER B M. Breaking through the tight junction barrier[J]. J Cell Biol, 1993, 123(6):1631-1633. doi: 10.1083/jcb.123.6.1631 [7] BAAS A F, KUIPERS J, VAN DER WEL N N, et al. Complete polarization of single intestinal epithelial cells upon activation of LKB1 by STRAD[J]. Cell, 2004, 116(3):457-466. doi: 10.1016/S0092-8674(04)00114-X [8] TSUKITA S, FURUSE M, ITOH M. Multifunctional strands in tight junctions[J]. Nat Rev Mol Cell Biol, 2001, 2(4):285-293. doi: 10.1038/35067088 [9] ROSE E C, ODLE J, BLIKSLAGER A T, et al. Probiotics, prebiotics and epithelial tight junctions: a promising approach to modulate intestinal barrier function[J]. Int J Mol Sci, 2021, 22(13):6729. doi: 10.3390/ijms22136729 [10] SUZUKI T. Regulation of the intestinal barrier by nutrients: the role of tight junctions[J]. Anim Sci J, 2020, 91(1):e13357. doi: 10.1111/asj.13357 [11] ADEWOLE D, AKINYEMI F. Gut microbiota dynamics, growth performance, and gut morphology in broiler chickens fed diets varying in energy density with or without bacitracin methylene disalicylate (BMD)[J]. Microorganisms, 2021, 9(4):787. doi: 10.3390/microorganisms9040787 [12] LI Y F, UDAYAKUMAR V, SATHUVAN M, et al. Effects of laminarin zwitterionic carboxylate and sulfonate on the intestinal barrier function and gut microbiota[J]. Carbohydr Polym, 2022, 278:118898. doi: 10.1016/j.carbpol.2021.118898 [13] YADAV M K, KUMARI I, SINGH B, et al. Probiotics, prebiotics and synbiotics: safe options for next-generation therapeutics[J]. Appl Microbiol Biotechnol, 2022, 106(2):505-521. doi: 10.1007/s00253-021-11646-8 [14] MERCADO-LUBO R, MCCORMICK B A. The interaction of gut microbes with host ABC transporters[J]. Gut Microbes, 2010, 1(5):301-306. doi: 10.4161/gmic.1.5.12925 [15] ZHAI Q X, FENG S S, ARJAN N, et al. A next generation probiotic, Akkermansia muciniphila[J]. Crit Rev Food Sci Nutr, 2019, 59(19):3227-3236. doi: 10.1080/10408398.2018.1517725 [16] GRAJEDA-IGLESIAS C, DURAND S, DAILLÈRE R, et al. Oral administration of Akkermansia muciniphila elevates systemic antiaging and anticancer metabolites[J]. Aging, 2021, 13(5):6375-6405. doi: 10.18632/aging.202739 [17] BEHROUZI A, VAZIRI F, RIAZI RAD F, et al. Comparative study of pathogenic and non-pathogenic Escherichia coli outer membrane vesicles and prediction of host-interactions with TLR signaling pathways[J]. BMC Res Notes, 2018, 11(1):539. doi: 10.1186/s13104-018-3648-3 [18] SAKSENA S, GOYAL S, RAHEJA G, et al. Upregulation of P-glycoprotein by probiotics in intestinal epithelial cells and in the dextran sulfate sodium model of colitis in mice[J]. Am J Physiol Gastrointest Liver Physiol, 2011, 300(6):G1115-G1123. doi: 10.1152/ajpgi.00027.2011 [19] DU F, HUANG R F, LIN D, et al. Resveratrol improves liver steatosis and insulin resistance in non-alcoholic fatty liver disease in association with the gut microbiota[J]. Front Microbiol, 2021, 12:611323. doi: 10.3389/fmicb.2021.611323 [20] LI W D, WANG Y X, SUN M, et al. The prebiotic-like effects of Coprinus comatus polysaccharides on gut microbiota in normal mice and those with acute alcoholic liver injury: a comparative study[J]. Evid Based Complementary Altern Med, 2020, 2020:2027570. [21] LI F T, YANG D, SONG F Y, et al. In vitro effects of ginseng and the seed of Zizyphus jujuba var. spinosa on gut microbiota of rats with spleen deficiency[J]. Chem Biodivers, 2020, 17(9):e2000199. doi: 10.1002/cbdv.202000199 [22] ZHANG Y B, LIU X Y, WANG Y F, et al. Antibacterial activity and mechanism of cinnamon essential oil against Escherichia coli and Staphylococcus aureus[J]. Food Contr, 2016, 59:282-289. doi: 10.1016/j.foodcont.2015.05.032 [23] LI Z X, ZHAO Y X, CHENG J L, et al. Integrated plasma metabolomics and gut microbiota analysis: the intervention effect of Jiawei Xiaoyao San on liver depression and spleen deficiency liver cancer rats[J]. Front Pharmacol, 2022, 13:906256. doi: 10.3389/fphar.2022.906256 [24] YANG Y N, CAO S J, XU W Y, et al. Dual modulation of gut bacteria and fungi manifests the gut-based anti-hyperlipidemic effect of Coptidis Rhizoma[J]. Biomed Pharmacother, 2022, 153:113542. doi: 10.1016/j.biopha.2022.113542 [25] TIAN Q M, WEI S M, SU H R, et al. Bactericidal activity of Gallic acid against multi-drug resistance Escherichia coli[J]. Microb Pathog, 2022, 173:105824. doi: 10.1016/j.micpath.2022.105824 [26] YANG J, QIAN D, JIANG S, et al. Identification of rutin deglycosylated metabolites produced by human intestinal bacteria using UPLC-Q-TOF/MS[J]. J Chromatogr B Analyt Technol Biomed Life Sci, 2012, 898:95-100. doi: 10.1007/s12272-017-0986-y [27] BAO Y, HAN X, LIU D, et al. Gut microbiota: the key to the treatment of metabolic syndrome in traditional Chinese medicine - a case study of diabetes and nonalcoholic fatty liver disease[J]. Front Immunol, 2022, 13:1072376. doi: 10.3389/fimmu.2022.1072376 [28] SHEN H, GAO X J, LI T, et al. Ginseng polysaccharides enhanced ginsenoside Rb1 and microbial metabolites exposure through enhancing intestinal absorption and affecting gut microbial metabolism[J]. J Ethnopharmacol, 2018, 216:47-56. doi: 10.1016/j.jep.2018.01.021 [29] GUO Y P, SHAO L, CHEN M Y, et al. In vivo metabolic profiles of Panax notoginseng saponins mediated by gut microbiota in rats[J]. J Agric Food Chem, 2020, 68(25):6835-6844. doi: 10.1021/acs.jafc.0c01857 [30] SONG R, XU L, XU F G, et al. Metabolic analysis of rhubarb extract by rat intestinal bacteria using liquid chromatography-tandem mass spectrometry[J]. Biomed Chromatogr, 2011, 25(3):417-426. doi: 10.1002/bmc.1467 [31] HUANG Z H, XU Y, WANG Q, et al. Metabolism and mutual biotransformations of anthraquinones and anthrones in rhubarb by human intestinal flora using UPLC-Q-TOF/MS[J]. J Chromatogr B Analyt Technol Biomed Life Sci, 2019, 1104:59-66. doi: 10.1016/j.jchromb.2018.10.008 [32] GOC J, LV M Z, BESSMAN N J, et al. Dysregulation of ILC3s unleashes progression and immunotherapy resistance in colon cancer[J]. Cell, 2021, 184(19): 5015-5030. [33] RACOVA Z, ANZENBACHEROVA E, PAPOUSKOVA B, et al. Metabolite profiling of natural substances in human: in vitro study from fecal bacteria to colon carcinoma cells (Caco-2)[J]. J Nutr Biochem, 2020, 85:108482. doi: 10.1016/j.jnutbio.2020.108482 [34] ENRIGHT E F, GOVINDARAJAN K, DARRER R, et al. Gut microbiota-mediated bile acid transformations alter the cellular response to multidrug resistant transporter substrates in vitro: focus on P-glycoprotein[J]. Mol Pharm, 2018, 15(12):5711-5727. doi: 10.1021/acs.molpharmaceut.8b00875 [35] TELBISZ Á, ÖZVEGY-LACZKA C, HEGEDŰS T, et al. Effects of the lipid environment, cholesterol and bile acids on the function of the purified and reconstituted human ABCG2 protein[J]. Biochem J, 2013, 450(2):387-395. doi: 10.1042/BJ20121485 [36] KELLY C J, ZHENG L, CAMPBELL E L, et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function[J]. Cell Host Microbe, 2015, 17(5):662-671. doi: 10.1016/j.chom.2015.03.005 [37] LI Y Y, JI X Y, WU H N, et al. Mechanisms of traditional Chinese medicine in modulating gut microbiota metabolites-mediated lipid metabolism[J]. J Ethnopharmacol, 2021, 278:114207. doi: 10.1016/j.jep.2021.114207 [38] CHEN M Y, LIAO Z Q, LU B Y, et al. Huang-Lian-Jie-du-decoction ameliorates hyperglycemia and insulin resistant in association with gut microbiota modulation[J]. Front Microbiol, 2018, 9:2380. doi: 10.3389/fmicb.2018.02380 [39] LOUIS P, HOLD G L, FLINT H J. The gut microbiota, bacterial metabolites and colorectal cancer[J]. Nat Rev Microbiol, 2014, 12(10):661-672. doi: 10.1038/nrmicro3344 [40] XIE X H, LIAO J B, AI Y L, et al. Pi-Dan-Jian-qing decoction ameliorates type 2 diabetes mellitus through regulating the gut microbiota and serum metabolism[J]. Front Cell Infect Microbiol, 2021, 11:748872. doi: 10.3389/fcimb.2021.748872 [41] KRAEHENBUHL J P, PRINGAULT E, NEUTRA M R. Review article: intestinal epithelia and barrier functions[J]. Aliment Pharmacol Ther, 1997, 11(s3):3-9. doi: 10.1111/j.1365-2036.1997.tb00803.x [42] GAO Z Z, LI Q W, WU X M, et al. New insights into the mecha-nisms of Chinese herbal products on diabetes: a focus on the “bacteria-mucosal immunity-inflammation-diabetes” axis[J]. J Immunol Res, 2017, 2017:1813086. [43] SHEN J, CHENG J Z, ZHU S G, et al. Regulating effect of baicalin on IKK/IKB/NF-kB signaling pathway and apoptosis-related proteins in rats with ulcerative colitis[J]. Int Immunopharmacol, 2019, 73:193-200. doi: 10.1016/j.intimp.2019.04.052 [44] YANG C C, DU Y, REN D Y, et al. Gut microbiota-dependent catabolites of tryptophan play a predominant role in the protective effects of turmeric polysaccharides against DSS-induced ulcerative colitis[J]. Food Funct, 2021, 12(20):9793-9807. doi: 10.1039/D1FO01468D [45] CHEN S Y, CHEN Z J, WANG Y, et al. Targeted delivery of Chinese herb pair-based berberine/tannin acid self-assemblies for the treatment of ulcerative colitis[J]. J Adv Res, 2022, 40:263-276. doi: 10.1016/j.jare.2021.11.017 [46] NIU C, HU X L, YUAN Z W, et al. Pulsatilla decoction improves DSS-induced colitis via modulation of fecal-bacteria-related short-chain fatty acids and intestinal barrier integrity[J]. J Ethnopharmacol, 2023, 300:115741. doi: 10.1016/j.jep.2022.115741 [47] GHOSH S, SINGH R, VANWINKLE Z M, et al. Microbial metabolite restricts 5-fluorouracil-resistant colonic tumor progression by sensitizing drug transporters via regulation of FOXO3-FOXM1 axis[J]. Theranostics, 2022, 12(12):5574-5595. doi: 10.7150/thno.70754 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 6332
- HTML全文浏览量: 4620
- PDF下载量: 205
- 被引次数: 0
 下载:
下载:
