

 下载:
下载:
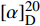
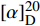
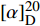
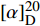
 下载:
下载:

-
麻黄碱是中药麻黄的主要有效成分,但它也可以引起中枢神经系统毒副作用。麻黄碱属于小分子生物碱,能够通过血脑屏障对神经细胞产生直接毒性作用 [1-4]。然而,有关麻黄碱神经毒性的作用机制并不完全清楚。脑源性神经营养因子(BDNF)已被证实参与学习、记忆等多种中枢神经系统的活动[5]。突触后致密蛋白是突触后膜胞质面聚集的一层均匀而致密的蛋白,其中PSD95是主要成分之一[6]。而synapsin1是维持突触前膜功能的重要成分之一[7]。BNDF、PSD95和synapsin1与神经元结构完整性和功能的完备性密切相关。本研究以PC12细胞为研究对象,考察麻黄碱的细胞毒性,同时检测BDNF、PSD95和synapsin1表达水平的变化,初步探讨麻黄碱引起细胞毒性的可能机制。
-
盐酸麻黄碱(批号:D03150702,赤峰艾克制药科技有限公司);BDNF抗体(批号:ab108319)、PSD95抗体(批号:ab18258)、synapsin1抗体(批号:ab64581)均购自英国Abcam公司。
-
PC12细胞购自中科院细胞库。
-
电子天平(德国Sartorious公司);孵育箱(Thermo公司);酶标仪(BioTek公司);Western blot设备(美国Bio-Rad公司)。
-
将培养瓶里的PC12细胞接种到96孔板上,每孔接种6000个细胞,用含1%FBS(胎牛血清)的RPMI 1640培养12 h后,吸弃原培养基,加入RPMI 1640基础培养基,1 h后分别加入不同浓度的麻黄碱,使其终浓度分别为0.25、0.5、1、2、4、8、16、20 mmol,继续培养24 h后,加入终浓度为0.5 mg/ml的MTT,继续孵育4 h,吸弃培养液,每孔加入150 μl的DMSO(二甲基亚砜),轻微震荡10 min,在酶标仪检测570 nm处的光密度(OD)值。
-
将培养瓶的PC12细胞接种到12孔板中,用含1%FBS的RPMI 1640培养12 h后,吸弃原培养基,加入RPMI 1640基础培养基1 h后分别加入不同浓度的麻黄碱,使其终浓度分别为10、500、1000 μmol,继续培养24 h后,在倒置显微镜下明场视野观察细胞形态变化。
-
将6孔板每孔接种105个PC12细胞,用含1%FBS的RPMI1640培养12 h,换上基础培养基RPMI1640,1h后分别加入不同浓度的麻黄碱,其终浓度分别为1、10、100、500、1000 μmol,继续培养24 h后,迅速将培养基吸弃,置于−30℃冷冻数小时,然后在冰上每孔加相应量的RIPA(含1%磷酸酶抑制剂和1%蛋白酶抑制剂)裂解液,4℃震荡30 min,将细胞裂解液收集到相应离心管里,4℃低温高速(12000 r/min)离心10 min。取出上清,蛋白浓度检测(BCA)试剂盒测定蛋白浓度后,加入相应量的5×的上样缓冲液和适量的RIPA裂解液,95℃加热10 min制样。制胶上样,样品80 V电泳结束后,400 mA电流90 min转移至PVDF(聚偏氟乙烯膜)后,5%脱脂奶粉室温封闭2 h,按分子量切出相应条带,分别孵育相应一抗过夜,洗涤后室温孵育相应二抗2 h,漂洗后,增强化学发光(ECL)法显影。
-
实验数据以
$ \bar x \pm s $ 表示。应用SPSS 20.0统计软件,采用单向方差分析法处理试验结果。首先进行Levene方差齐性检验,方差齐性且整体比较组间有显著性差异时,采用LSD法进行多重比较,方差分析不显著时不进行多重比较;若方差不齐,则作Welch检验,方差分析有显著性差异时,采用Dunnett’s T3法进行多重比较,方差分析不显著时不进行多重比较。 -
MTT结果如图1所示,麻黄碱在一定范围内对PC12细胞有一定的毒性,经拟合计算在PC12细胞中麻黄碱的IC25值为0.536 mmol,IC50值为2.8 mmol。
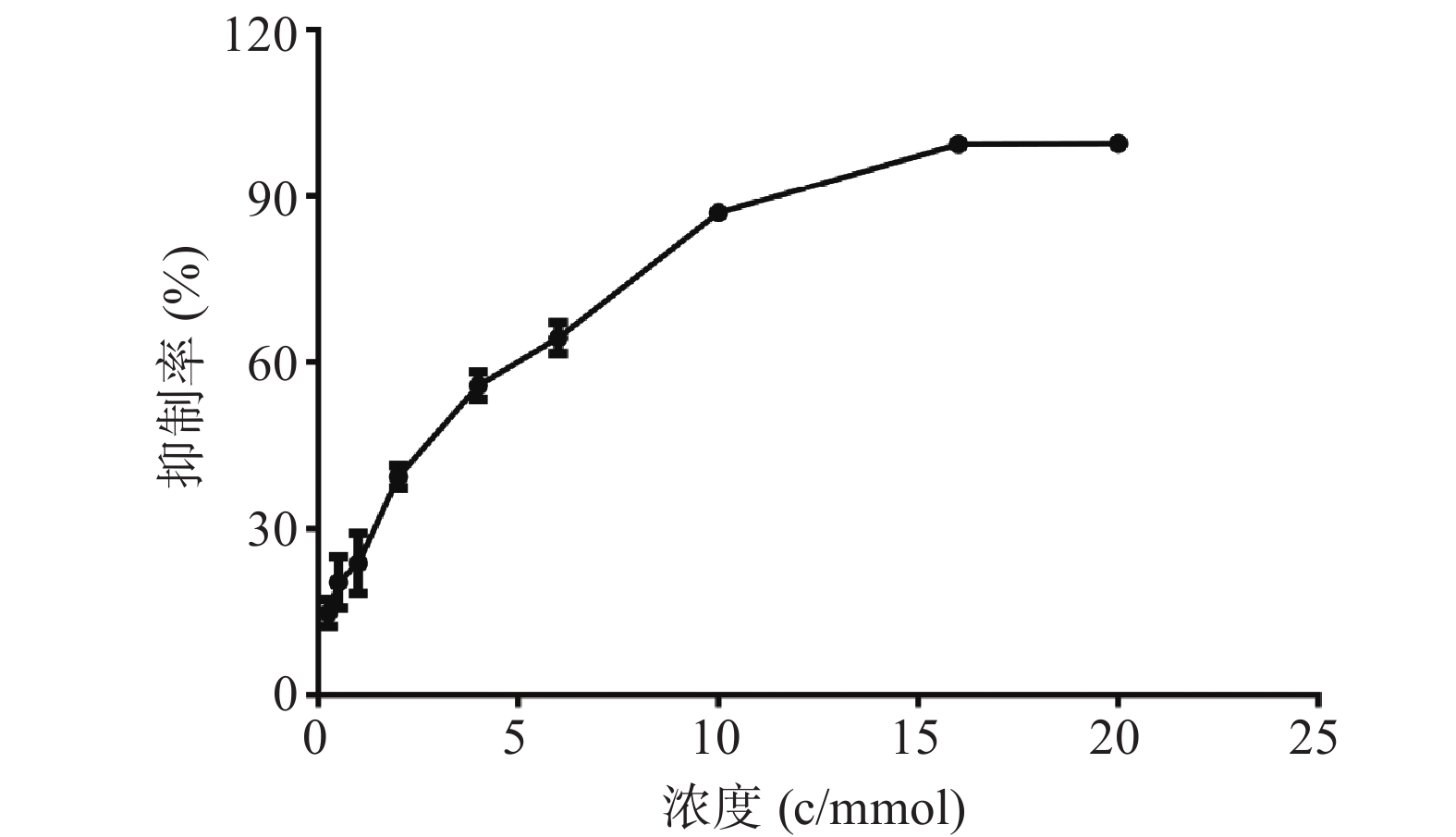
图 1 麻黄碱细胞活性试验(MTT法)
-
PC12细胞形态学变化如图2所示,在20倍镜下观察,随着麻黄碱浓度的升高细胞数量与空白对照组相比逐渐减少,细胞突触数量也随之减少。1000 μmol浓度处理条件下,轴突长度显著减短,细胞体边缘模糊,细胞体积显著变小。
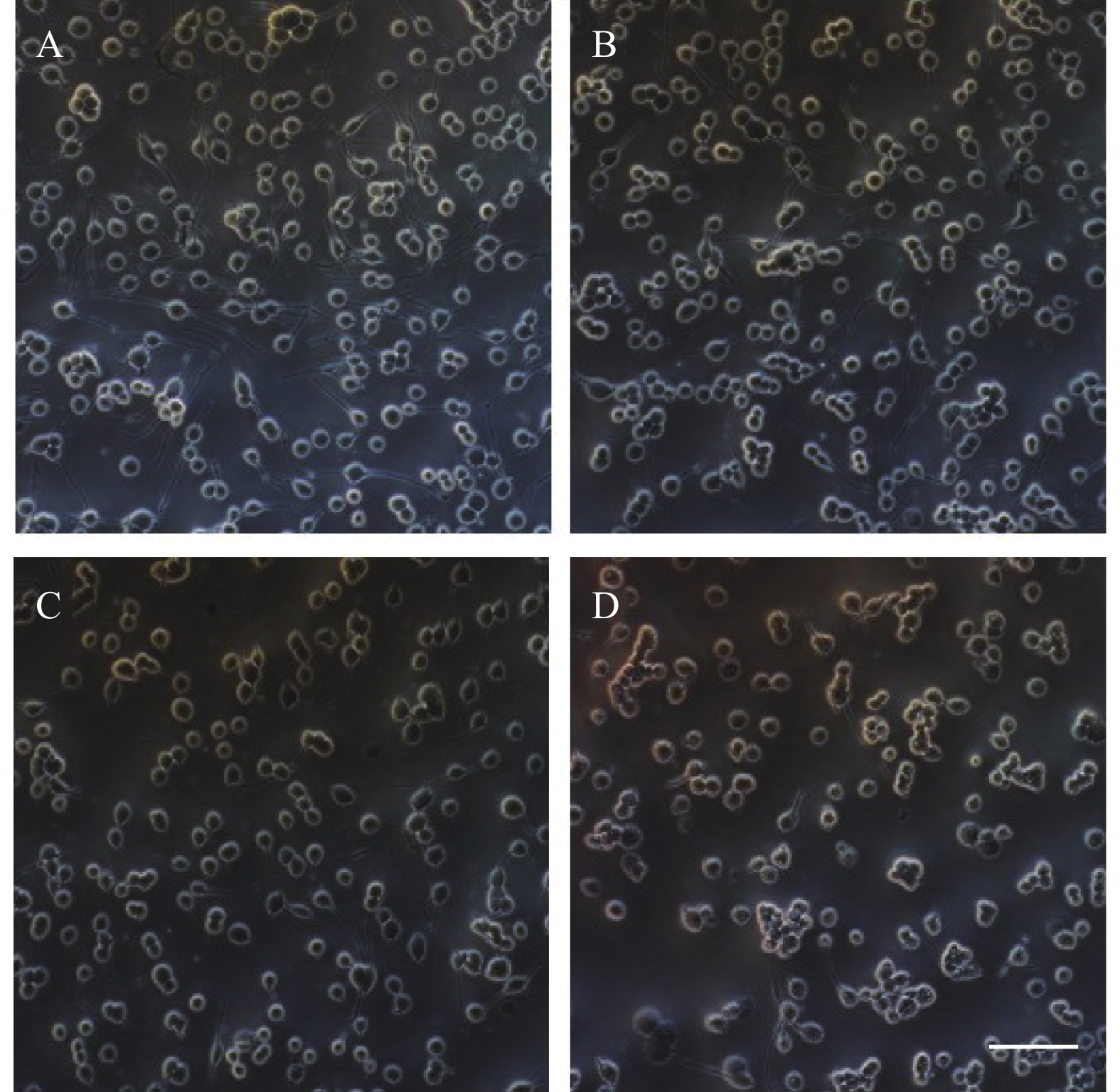
图 2 麻黄碱对PC12细胞形态变化的影响
-
图3结果显示,随着麻黄碱剂量的增加,PC12细胞中BDNF的蛋白表达量增加。当麻黄碱浓度为500 μmol时,BDNF的表达量约为空白对照组的1.58倍,有统计学差异(P<0.05);而当麻黄碱浓度高达 1000 μmol时,BNDF为对照组的1.91倍(P<0.001)。说明该浓度范围内麻黄碱可以浓度依赖性的引起PC12细胞中BDNF表达升高。
图 3 麻黄碱对BDNF表达水平变化的影响
-
图4结果显示,随着麻黄碱剂量的增加,PC12细胞中PSD95的表达增加。麻黄碱浓度为500 μmol时,PSD95的蛋白表达水平约为空白对照组的1.38倍,有统计学差异(P<0.05);而当麻黄碱浓度达到1000 μmol时,PSD95表达量约上升为对照组的1.52倍(P<0.001)。说明该浓度范围内麻黄碱可以浓度依赖性的引起PC12细胞中PSD95表达升高。
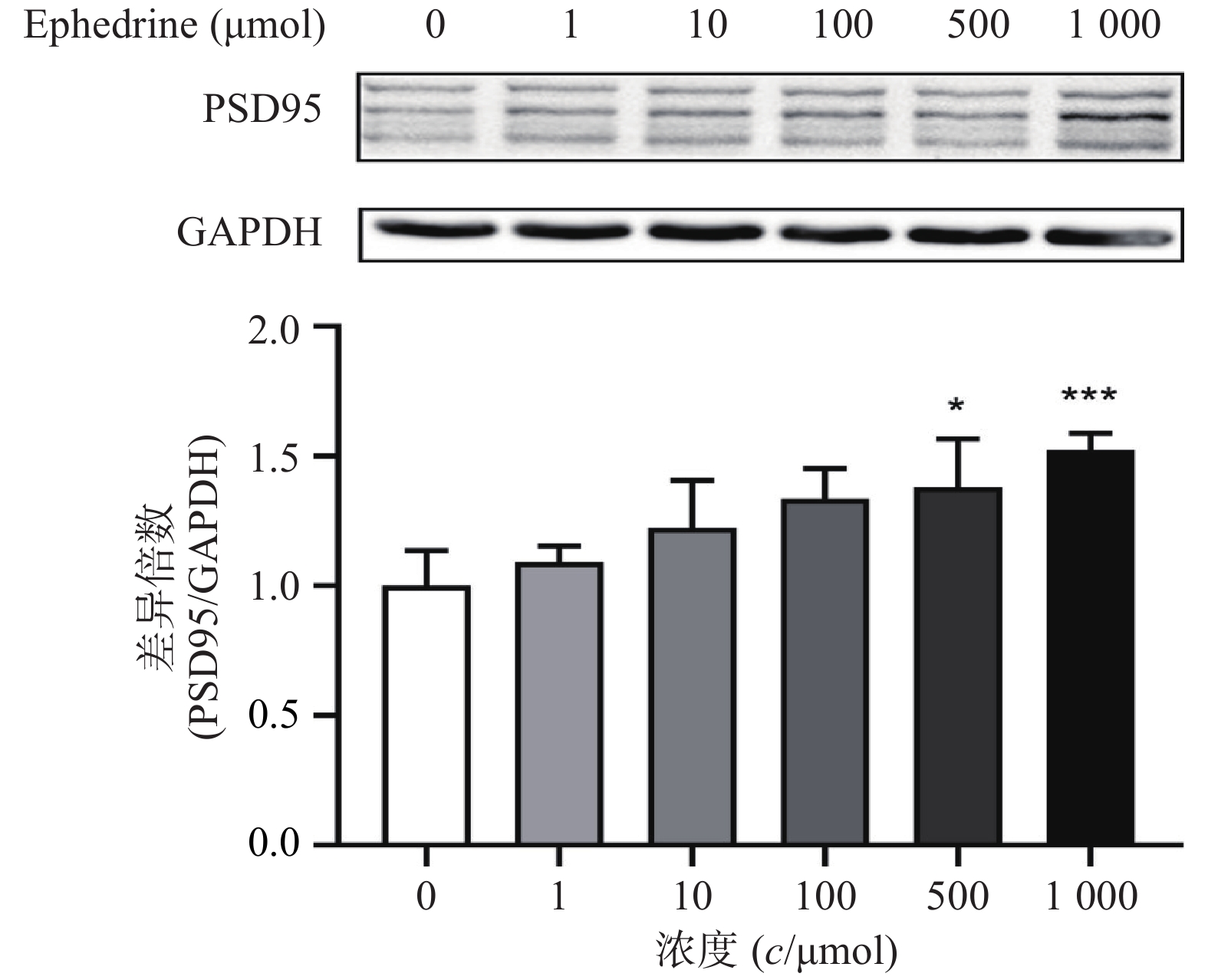
图 4 麻黄碱对PSD95表达水平变化的影响
-
图5结果显示,随着麻黄碱剂量的增加,PC12细胞中synapsin1的表达量有下降的趋势。其中,麻黄碱浓度为500 μmol时,synapsin1的蛋白表达水平约为空白对照组的0.65,具有统计学差异(P<0.05);而当麻黄碱浓度达到1000 μmol时,synapsin1表达量下降至对照组的0.53(P<0.001)。说明麻黄碱在该浓度范围内可以引起PC12细胞中synapsin1表达降低。
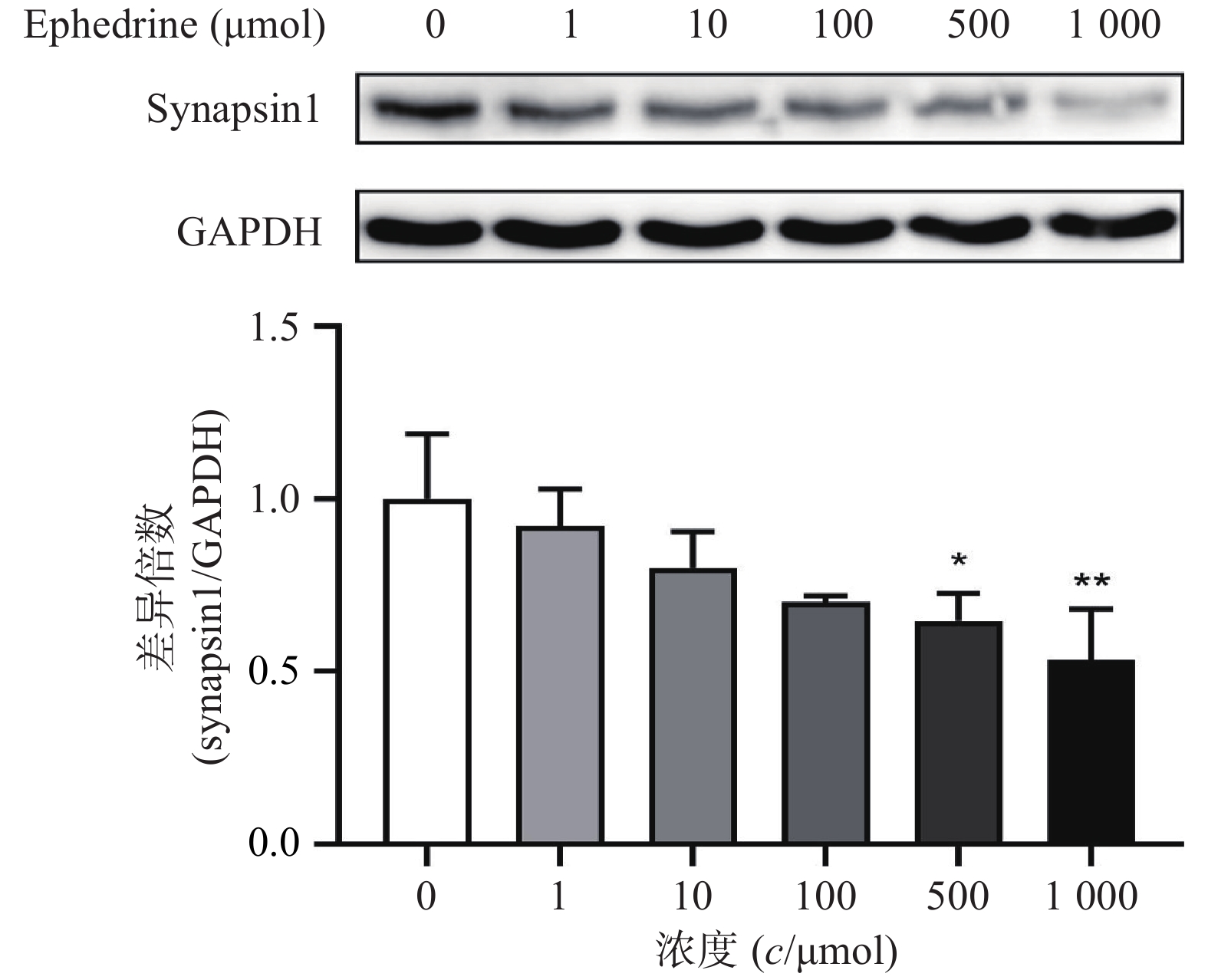
图 5 麻黄碱对synapsin1表达水平变化的影响
-
麻黄历来被中医冠以“解表第一要药”,是治疗外感风寒表实证的重要中药,主要用于风寒感冒,胸闷喘咳,风水浮肿等,麻黄汤就是以麻黄为君药的治疗表寒实证的代表名方。然而前期研究发现,给予大鼠连续灌胃麻黄,可以造成大鼠不同脑区损伤,其中对额叶皮层影响较大,HE染色结果甚至观察到神经元凋亡早期的形态学改变 [8-9]。已经证实麻黄中对中枢神经系统有兴奋毒性作用的主要成分是麻黄碱。麻黄碱的神经毒性可能与升高兴奋性谷氨酸水平有关,过量的兴奋性谷氨酸能引起多巴胺神经末梢的破坏和额叶皮层、丘脑、边缘系统的神经变性。为了明确麻黄引起中枢神经系统毒性的机制,本研究中将麻黄碱单体作为研究对象,考察其在体外细胞模型中的毒性作用。
PC12细胞源于鼠嗜铬细胞瘤,在结构上和功能上与神经元有高度相似的地方,因其可重复性好,被广泛应用于中枢神经系统的研究。本研究首先考察了不同浓度麻黄碱对PC12细胞存活率的影响,确定了麻黄碱引起细胞毒性的浓度范围。在此基础上,分别考察不影响细胞存活率情况下(IC25值以下浓度)及显著影响细胞存活率情况下(IC25值以上浓度),麻黄碱对中枢神经系统中相关功能蛋白的影响。
BDNF是神经营养因子家族的肽生长因子的一员。在体实验发现,BDNF在大脑海马的CA1区可通过促进兴奋性突触的发育来增强突触强度。同时,BDNF可以在皮层神经元中增加介导中枢神经中快速兴奋性突触传递的AMPA受体的表达,提高动物兴奋性[10-13]。而外源性的BDNF则可以通过增强突触间的传递,增强海马神经元的兴奋性 [14]。以上研究表明,BDNF与神经细胞的兴奋性密切相关。PSD95定位于突触后膜,是高电子密度的半圆形区域形成的突触后致密区的主要组成部分之一。研究发现在海马神经元中过表达PSD95可以促进成熟谷氨酸能突触以及谷氨酸受体的活性 [6,15]。PSD95链接了兴奋性受体NMDAR(N-甲基-D-天冬氨酸受体)和下游信号分子,并且在神经元成熟以及突触的形成中发挥着重要作用[16]。尽管在多数神经退行性病变中,提高BDNF和PSD95的表达水平可以显著改善行为学,但是也有研究表明在生理状态下,神经元的过度激活会引起神经元的功能的减退甚至导致神经损伤。由于BDNF和PSD95与神经元兴奋性关系密切,在前期的行为学实验中,表明麻黄碱灌胃给药会引起大鼠自主活动的增加,并激活神经元,故我们在考察麻黄碱产生细胞毒性的机制时,检测了BDNF和PSD95的蛋白表达水平。结果显示,随着麻黄碱浓度的增加,BDNF和PSD95的表达水平浓度依赖性地升高,当浓度达到500 μmol时差异具统计学意义(P<0.05),说明麻黄碱可以引起体外神经细胞模型的兴奋状态。
与此同时,通过观察细胞形态变化,发现一定剂量的麻黄碱会引起PC12细胞突触数量下降和长度减短,说明麻黄碱会引起突触丢失。文献研究表明,synapsin1是突触的重要标志物,可以反应突触形态上的完整性和功能[7]。由此,我们在考察麻黄碱对PC12细胞形态影响的同时,采用Western blot检测了synapsin1的表达水平。梯度浓度麻黄碱处理PC12细胞后synapsin1的表达变化结果表明随着麻黄碱处理浓度的增加,synapsin1的表达水平降低,当浓度达到500 μmol时差异具统计学意义(P<0.05),说明麻黄碱有一定的细胞毒性的同时,也可以引起神经细胞的突触毒性,突触蛋白的丢失可能也是引起神经细胞毒性的原因之一。
综上所述,本实验结果表明麻黄碱可以对PC12细胞产生毒性。与正常细胞相比,麻黄碱处理组的细胞在形态学上发生了明显的变化,BDNF和PSD95等与兴奋性相关的蛋白表达出现了明显升高,而与突触功能有关的蛋白synapsin1表达出现了明显下降,说明麻黄碱产生中枢神经系统毒副作用的机制与BDNF、PSD95及synapsin1的表达变化有关。
Effects of ephedrine on the expression levels of BDNF, PSD95 and synapsin1 in PC12 cells
-
摘要:
目的 考察麻黄碱对PC12细胞内的脑源性神经营养因子(BDNF)、突触后致密蛋白95(PSD95)和神经突触素1(synapsin1)表达水平的影响,探讨麻黄碱对PC12细胞毒性的作用机制。 方法 通过不同浓度的麻黄碱处理PC12细胞后,采用噻唑蓝试剂(MTT)法测定细胞存活率;采用倒置显微镜观察细胞的形态学变化;采用蛋白印迹(Western blot)法检测BDNF、PSD95和synapsin1的蛋白表达水平。 结果 麻黄碱呈浓度依赖性降低PC12细胞活力,其引起PC12细胞死亡的IC25和IC50分别为0.536 mmol和2.8 mmol。随着麻黄碱浓度的升高,PC12细胞体积变小,边界模糊,突触数减少,轴突长度减短;BDNF和PSD95的表达水平明显升高;synapsin1的表达水平有所降低。 结论 麻黄碱对PC12细胞的毒性作用机制可能与影响BDNF、PSD95和synapsin1的表达水平有关。 Abstract:Objective To investigate the effects of ephedrine on the expression levels of brain-derived neurotropic factor (BDNF) and postsynaptic density protein 95 (PSD95) and synapsin1 in PC12 cells, and to explore the mechanism of ephedrine cytotoxicity on PC12. Methods After PC12 cells were treated with different concentration of ephedrine, the cell survival rate was measured by the methylthiazolyldiphenyl-tetrazolium bromide (MTT) assay. The morphology changes of PC12 cells were observed by an inverted microscope. Western blot was used to detect the protein expression levels of BDNF, PSD95 and synapsin1 in PC12 cells. Results Ephedrine decreased the viability of PC12 cell in a concentration-dependent manner,with an IC25 and IC50 of 0.536 mmol and 2.8 mmol, respectively, for PC12 cell death. As ephedrine concentration increased, PC12 cells became smaller in size, with blurred boundary blurred, reduced synapses and shorter axon lengths. The expression levels of BDNF and PSD95 increased significantly. Meanwhile the expression level of synapsin1 decreased. Conclusion The mechanism of ephedrine cytotoxicity on PC12 may be related to the expression levels of BDNF, PSD95 and synapsin1. -
海绵是具有代表性的海洋生物,其共附生微生物也是近年来研究的热点。在海洋高盐、高压、低温、寡营养的生存环境下,海绵共附生微生物能够产生结构新颖、生物活性良好的次级代谢产物。其中海绵共附生真菌是海绵化学多样性的重要来源[1]。
曲霉属 (Aspergillus sp)真菌分布广泛而且研究丰富。海洋曲霉属真菌的次级代谢产物主要包括聚酮类[2]、生物碱类[3]、肽类[4]、萜类[5]等化合物,具有抗肿瘤[6]、抗菌[7]、抗病毒[4]等生物活性。本课题的土曲霉(Aspergillus terreus)是从我国南海西沙永兴岛海域的棕色扁海绵Phakellia fusca中分离得到的,属于散囊菌目(Eurotiales)发菌科(Tri-chocomaceaez)的一种真菌,在海洋动植物和陆地植物中均有分布。该菌的次级代谢产物具有多样性,包括生物碱类化合物[8]、丁烯酸内酯类化合物[9]、萜类化合物[10]、环肽类化合物[11]等。本文采用硅胶柱色谱、Sephadex LH-20凝胶柱色谱、高效液相色谱等多种分离方法从土曲霉Aspergillus terreus中共分离得到8个单体化合物。通过理化常数测定、波谱数据分析等方法确定了化合物的结构。化合物1~8的结构见图1。
1. 材料和方法
1.1 样品
菌株来源于棕色扁海绵Phakellia fusca,由上海交通大学海洋药物研究中心鉴定为Aspergillus terreus,菌株保存在上海交通大学医学院附属仁济医院药学部海洋药物研究中心(菌株编号152805)。
1.2 仪器与试剂
Agilent 600核磁共振波谱仪(美国 Agilent 公司);Waters高效液相色谱仪(美国Waters公司);XBridge C18半制备型液相色谱柱(10 mm×250 mm,5 μm);快速制备色谱仪(法国Interchim公司);OSB-2100旋转蒸发仪(日本EYELA 公司);振荡培养箱(上海知楚)。薄层硅胶、200~300目柱色谱用硅胶(青岛海洋化工厂);Sephadex LH-20凝胶(瑞典GE Healthcare公司);色谱纯试剂(天津康科德科技有限公司);其他分析纯有机试剂(上海化学试剂公司);氘代试剂(剑桥同位素实验室)。
1.3 发酵与萃取
取Aspergillus terreus单菌落接种到装有100 ml PDB培养液的250 ml三角瓶中,28 ℃,220 r/min震荡培养3 d,以该发酵液10%的接种量接到装有500 ml的真菌2号培养液(甘露醇20 g,麦芽糖20 g,CaCO3 15 g,葡萄糖10 g,谷氨酸钠10 g,酵母提取物3 g,玉米浆1 g,KH2PO4 0.5 g,MgSO4·7H2O 0.3 g,海盐30 g,蒸馏水1 L)的1 L三角瓶中,28 ℃,220 r/min震荡培养10 d,获得菌株的发酵物。收集发酵液24 L,用等体积的乙酸乙酯萃取3次,浓缩后得到乙酸乙酯相浸膏9.3 g。
1.4 提取分离
乙酸乙酯相浸膏首先经Sephadex LH-20凝胶柱色谱分离,以二氯甲烷-甲醇(体积比为1∶1)作为溶剂进行洗脱,得到组分Fr.1~Fr.4。组分Fr.2经硅胶柱色谱(石油醚:丙酮 = 100∶1~0∶100)分离得到组分Fr.2-1~Fr.2-9。组分Fr.2-5经反相中压柱色谱分离得到8个亚组分,其中Fr.2-5d经重结晶得到化合物3 (2.5 mg)。组分Fr.2-6经LH-20凝胶柱色谱和反相半制备HPLC(38%乙腈-水)分离得到化合物1 (3.5 mg, tR = 21.0 min)。化合物2 (3.5 mg, tR = 13.0 min)由组分Fr.2-7经反相半制备HPLC,以33%乙腈-水为流动相等梯度洗脱得到。组分Fr.2-8以乙腈-水 (体积比10∶90~100∶0)为流动相,经反相中压柱色谱和反相半制备HPLC(20%乙腈-水)分离得到化合物4 (2.0 mg, tR=30.0 min)、 化合物5 (4.0 mg, tR=28.0 min)和化合物6 (9.0 mg, tR=14.0 min)。Fr.3经过硅胶柱色谱分离得到7个组分,其中Fr.3-3经反相半制备HPLC进一步纯化得到化合物7 (1.7 mg, tR=12.0 min)。组分Fr.3-4以20%~100%的乙腈-水为流动相,经反相中压柱色谱和反相半制备HPLC(15%乙腈-水)分离得到化合物8 (18.0 mg, tR = 8.0 min)。
2. 结构鉴定
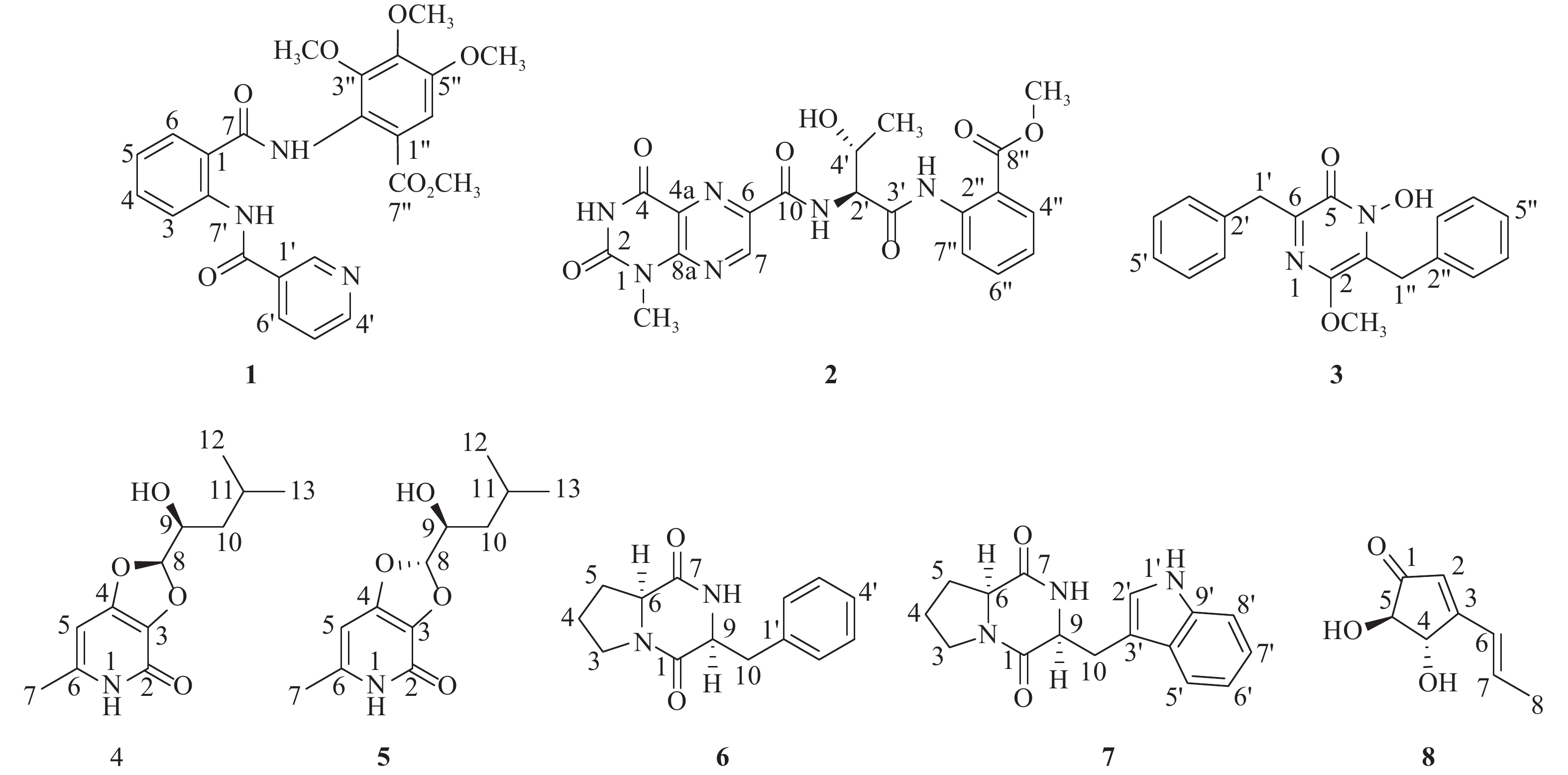
化合物1为黄色粉末(甲醇),硫酸/香草醛显色为黄色,ESIMS给出的分子离子峰[M+H]+m/z 466.15。1H NMR (600 MHz, CDCl3)中,δH 12.23 (1H, s)为氨基质子信号;一组邻位二取代的苯环质子信号δH 8.82 (1H, dd, J=8.5, 0.8 Hz, H-3), 7.89 (1H, dd, J=7.9, 1.3 Hz, H-6), 7.60 (1H, td, J=8.5, 1.3 Hz, H-4), 7.22 (1H, m, H-5),芳香质子信号δH 9.21 (1H, brs, H-9), 8.70 (1H, d, J=4.5 Hz, H-1′), 8.25 (1H, dt, J=8.0, 2.2 Hz, H-3′), 7.36 (1H, dd, J=8.0, 4.5 Hz, H-2′),提示3-取代吡啶环的存在;1个芳香质子信号δH 7.27 (1H, s, H-10′);4个甲氧基质子信号δH 3.97 (3H, s, 4″-OCH3), 3.91 (3H, s, 3″-OCH3), 3.90 (3H, s, 5″-OCH3), 3.82 (3H, s, 7″-OCH3)。13C NMR (150 MHz, CDCl3)共显示24个碳信号,结合DEPT谱,推断δC 168.2, 167.2, 164.0为羰基碳信号;17个芳香碳信号;δC 61.3, 61.3, 56.5, 52.7为4个甲氧基碳信号。碳信号归属为:δC 168.2 (C-7)、167.2 (C-7′′)、164.0 (C-7′)、152.6 (C-4′)、151.5 (C-5′′)、149.3 (C-2′)、148.8 (C-3′′)、146.9 (C-4′′)、140.4 (C-2)、135.2 (C-6′)、133.6 (C-4)、130.3 (C-1′)、127.9 (C-6)、125.8 (C-2′′)、123.8 (C-5)、123.6 (C-5′)、121.8 (C-3)、120.4 (C-1′′)、119.0 (C-1)、108.8 (C-6′′)、61.3 (3″-OCH3)、61.3 (4″-OCH3)、56.5 (5″-OCH3)、52.7 (7″-OCH3)。该化合物核磁数据与参考文献[11]对照基本一致,确定化合物为methyl-3,4,5-trimethoxy-2-(2-(nicotinamido)benzamido) benzoate。
化合物2为黄色粉末(甲醇),ESIMS给出的分子离子峰[M+H]+m/z 457.14。1H NMR (600 MHz, DMSO-d6)中,δH 12.19 (1H, s, 3-NH), 11.10 (1H, s, 1′′-NH), 8.52 (1H, d, J = 8.1 Hz, 1′-NH)为氨基质子信号;1个芳香质子单峰信号δH 9.29 (1H, s, H-7);一组邻位二取代的苯环质子信号δH 8.44 (1H, d, J = 8.5 Hz, H-7′′), 7.92 (1H, dd, J = 7.9, 1.5 Hz, H-4′′), 7.63 (1H, td, J = 7.9, 1.5 Hz, H-6′′), 7.20 (1H, td, J = 7.6, 1.5 Hz, H-5′′);2个相邻的连接杂原子的次甲基质子信号δH 4.55 (1H, dd, J = 8.1, 2.9 Hz, H-2′), 4.41 (1H, m, H-4′);3个甲基质子信号δH 3.70 (3H, s, H-9′′), 3.52 (3H, s, H-9), 1.19 (3H, d, J = 6.4 Hz, H-5′)。13C NMR (150 MHz, DMSO-d6)共显示20个碳信号,结合DEPT谱,推断δC 168.8, 167.3, 162.7, 159.5, 150.1为羰基碳信号;10个芳香碳信号;δC 65.9, 59.8为2个连杂原子的次甲基碳信号;δC 52.4, 28.6, 20.5为3个甲基碳信号,结合氢谱信号,确定有一个甲氧基和一个氮甲基。碳信号归属为:δC 168.8 (C-3′)、167.3 (C-8″)、162.7 (C-10)、159.5 (C-4)、151.2 (C-8a)、150.1 (C-2)、146.3 (C-7)、139.3 (C-2′′)、138.2 (C-6)、134.2 (C-6′′)、130.7 (C-4′′)、127.2 (C-4a)、123.4 (C-5′′)、120.7 (C-7′′)、117.1 (C-3′′)、65.9 (C-4′)、59.8 (C-2′)、52.4 (C-9″)、28.6 (C-9)、20.5 (C-5′)。该化合物的比旋光值为
$[\alpha]_{\rm{D}}^{20} $ +98 (c 0.1, MeOH)。该核磁数据与参考文献[12]对照基本一致,确定该化合物为terrelumamide A。化合物3为白色结晶(甲醇),ESIMS给出的分子离子峰[M+H]+m/z 323.13。1H-NMR (600 MHz, CDCl3)中,δH 7.2-7.5 (10H, m, H-3′-H-7′, H-3′′-H-7′′)为10个芳香质子信号,提示存在2个单取代苯基;2个亚甲基质子信号δH 4.20 (2H, brs, H-1′′), 3.94 (2H, brs, H-1′);1个甲氧基质子信号δH 3.92 (3H, s, 2-OCH3)。13C-NMR (150 MHz, CDCl3)共显示19个碳信号,结合DEPT谱推断δC 158.2为羰基碳信号;12个芳香碳信号;δC 34.0, 30.4为2个亚甲基碳信号,提示结构中存在两个苄基基团;δC 61.8为甲基碳信号;δC 144.2, 140.6, 129.4为3个烯碳信号。碳信号归属为:δC 158.2 (C-5), 144.2 (C-6), 140.6 (C-2), 136.5 (C-2′′), 135.6 (C-1′), 129.6 (C-3′, 7′), 129.4 (C-3, 3′′, 7′′), 128.6 (C-4′, 6′), 127.8 (C-4′′, 6′′), 126.9 (C-5′, 5′′), 61.8 (2-OCH3), 34.0 (C-1′′), 30.4 (C-1′)。该化合物核磁数据与参考文献[13]对照基本一致,确定化合物为emeheterone。
化合物4为黄色粉末(甲醇),硫酸/香草醛显色为紫色,ESIMS给出的分子离子峰[M+H]+m/z 240.12。1H NMR (600 MHz, CD3OD)中,给出1个芳香质子信号δH 6.13 (1H, d, J = 0.7 Hz, H-5);3个次甲基氢信号δH 6.07 (1H, d, J = 3.0 Hz, H-8), 3.89 (1H, dt, J = 10.5, 3.0 Hz, H-9), 1.90 (1H, m, H-11);1个亚甲基质子信号δH 1.58 (1H, ddd, J = 12.2, 10.5, 4.6 Hz, H-10), 1.36 (1H, ddd, J = 12.2, 10.5, 3.0 Hz, H-10);3个甲基质子信号δH 2.28 (3H, s, H-7), 0.99 (3H, d, J = 6.7 Hz, H-13), 0.96 (3H, d, J = 6.7 Hz, H-12)。13C NMR (150 MHz, CD3OD)共显示12个碳信号,结合DEPT谱推断δC 155.0为羰基碳信号;4个芳香碳信号;δC 115.8, 70.5, 25.2为3个次甲基脂肪碳信号,结合对应的氢信号提示结构中存在1个缩醛碳信号和一个连氧次甲基碳信号;δC 40.4为亚甲基碳信号;δC 24.0, 21.8, 18.8为3个甲基碳信号。碳谱信号归属为:δC 157.9 (C-4)、155.0 (C-2)、143.5 (C-6)、132.7 (C-3)、115.8 (C-8)、95.0 (C-5)、70.5 (C-9)、40.4 (C-10)、25.2 (C-11)、24.0 (C-12)、21.8 (C-13)、18.8 (C-7)。该化合物的ECD曲线显示在217 nm处有负的Cotton 效应(Δε −5.86),其核磁和ECD数据与参考文献[14]对照基本一致,最终确定该化合物为(8R, 9S)-dihydroisoflavipucine。
化合物5为黄色结晶(甲醇),硫酸/香草醛显色为紫色,ESIMS给出的分子离子峰[M+H]+m/z 240.12。1H NMR (600 MHz, CD3OD)中,给出1个芳香质子信号δH 6.13 (1H, d, J = 0.7 Hz, H-5);3个次甲基氢信号δH 6.06 (1H, d, J = 3.0 Hz, H-8), 3.90 (1H, dt, J = 10.5, 3.0 Hz, H-9), 1.90 (1H, m, H-11);1组亚甲基质子信号δH 1.56 (1H, ddd, J = 12.3, 10.5, 4.6 Hz, H-10), 1.36 (1H, ddd, J = 12.3, 10.5, 3.0 Hz, H-10);3个甲基质子信号δH 2.28 (3H, s, H-7), 0.99 (3H, d, J = 6.6 Hz, H-13), 0.95 (3H, d, J = 6.6 Hz, H-12)。13C NMR (150 MHz, CD3OD)共显示12个碳信号,结合DEPT谱推断δC 155.0为羰基碳信号;4个芳香碳信号;δC 115.8, 70.5, 25.2为3个次甲基碳信号,结合对应的氢信号提示结构中存在1个次甲二氧基碳信号和一个连氧次甲基碳信号;δC 40.5为亚甲基碳信号;δC 24.0, 21.8, 18.8为3个甲基碳信号。碳信号归属为:δC 157.8 (C-4)、155.0 (C-2)、143.4 (C-6)、132.8 (C-3)、115.8 (C-8)、95.1 (C-5)、70.5 (C-9)、40.5 (C-10)、25.2 (C-11)、24.0 (C-12)、21.8 (C-13)、18.8 (C-7)。该化合物的核磁数据与化合物4对比基本一致,ECD曲线显示在217 nm处有正的Cotton 效应(Δε +25.34),提示为化合物4的差向异构体。将此化合物的核磁和ECD数据与参考文献[14]对照基本一致,最终确定化合物为(8S, 9S)-dihydroisoflavipucine。
化合物6为黄色粉末(甲醇),硫酸/香草醛溶液无明显显色,ESIMS给出的分子离子峰[M+H]+m/z 245.12。1H NMR (600 MHz, CDCl3)中,给出1组单取代的苯环芳香质子信号δH 7.32 (2H, t, J = 7.5 Hz, H-5′), 7.26 (1H, t, J = 7.5 Hz, H-4′), 7.20 (2H, d, J = 7.5 Hz, H-6′);2个次甲基氢信号δH 4.25 (1H, dd, J=10.5, 2.9 Hz, H-9), 4.04 (1H, t, J = 7.8 Hz, H-6);4组亚甲基质子信号δH 3.65-3.50 (2H, m, H-3); 3.65-3.50 (1H, m, H-10), 2.76 (1H, dd, J=14.5, 10.5 Hz, H-10); 2.30 (1H, m, H-5), 1.88 (1H,m, H-5); 1.98 (2H, m, H-4)。13C NMR (150 MHz, CDCl3)共显示14个碳信号,结合DEPT谱推断δC 169.6, 165.3为酰胺羰基碳信号;6个芳香碳信号;δC 59.3, 56.4为2个连氮次甲基碳信号;δC 45.6, 37.0, 28.5, 22.7为4个亚甲基碳信号,提示结构中存在苯丙氨酸和脯氨酸片段。碳信号归属为:δC 169.6 (C-7)、165.3 (C-1)、136.1 (C-1′)、129.4 (C-2′)、129.4 (C-6′)、129.3 (C-3′)、129.3 (C-5′)、127.7 (C-4′)、59.3 (C-6)、56.4 (C-9)、45.6 (C-3)、37.0 (C-10)、28.5 (C-5)、22.7 (C-4)。该化合物的比旋光值为
$[\alpha]_{\rm{D}}^{20} $ -47 (c 0.1, MeOH),将核磁数据与参考文献[15]对照基本一致,最终确定化合物为cyclo(S-Pro-S-Phe)。化合物7为浅黄色粉末(甲醇),硫酸/香草醛显色不明显,ESIMS给出的分子离子峰[M+H]+m/z 284.13。1H NMR (600 MHz, DMSO-d6)中给出2个氨基质子信号δH 10.83 (1H, s, H-1′), 7.71 (1H, s, H-8);1组邻二取代的苯环芳香质子信号δH 7.54 (1H, d, J = 8.0 Hz, H-5′), 7.30 (1H, d, J = 8.0 Hz, H-8′), 7.03 (1H, t, J = 7.3 Hz, H-7′), 6.94 (1H, t, J = 7.3 Hz, H-6′);1个芳香质子单峰信号δH 7.16 (1H, s, H-2′);2个次甲基氢信号δH 4.28 (1H, t, J = 5.0 Hz, H-9), 4.04 (1H, t, J = 8.5 Hz, H-6);4组亚甲基质子信号δH 3.36 (1H, m, H-3), 3.23 (1H, m, H-10), 3.21(1H, m, H-3), 3.05 (1H, m, H-10), 1.95 (1H, m, H-5), 1.66 (1H, m, H-4), 1.59 (1H, m, H-4), 1.36 (1H, m, H-5)。13C NMR (150 MHz, DMSO-d6)共显示16个碳信号,结合DEPT谱推断δC 169.0, 165.5为酰胺羰基碳信号;8个芳香碳信号;δC 58.4, 55.2为2个连氮次甲基碳信号;δC 44.6, 27.7, 25.8, 21.8为4个亚甲基碳信号。碳信号归属为:δC 169.0 (C-7)、165.5 (C-1)、136.0 (C-9′)、127.3 (C-4′)、124.4 (C-2′)、120.8 (C-7′)、118.6 (C-5′)、118.2 (C-6′)、111.2 (C-8′)、109.3 (C-3′)、58.4 (C-6)、55.2 (C-9)、44.6 (C-3)、27.7 (C-5)、25.8 (C-10)、21.8 (C-4)。将核磁数据与化合物6对比,化合物7中吲哚基取代了化合物6中的苯基。该化合物的比旋光值为
$[\alpha]_{\rm{D}}^{20} $ -90 (c 0.1, MeOH),将该核磁数据与参考文献[16]对照基本一致,最终确定化合物为brevianamide F。化合物8为棕黄色油状(甲醇),ESIMS给出的分子离子峰[M+Na]+m/z 177.06。1H NMR (600 MHz, DMSO-d6)中,给出3个烯氢信号δH 6.72 (1H, m, H-7), 6.37 (1H, d, J = 15.8 Hz, H-6), 6.00 (1H, s, H-2),其中一对为反式烯氢;2个羟基信号δH 5.80 (1H, s, 5-OH), 5.68 (1H, s, 4-OH);2个连氧次甲基质子信号δH 4.50 (1H, m, H-4), 3.89 (1H, m, H-5);1个甲基质子信号δH 1.88 (3H, d, J = 6.3 Hz, H-8)。13C NMR (150 MHz, DMSO-d6)共显示8个碳信号,结合DEPT谱,推断δC 203.7为酮羰基碳信号;4个双键碳信号;δC 80.8, 76.4为2个连氧次甲基碳信号;δC 19.1为甲基碳信号。碳信号归属为:δC 203.7 (C-1)、168.5 (C-3)、139.4 (C-7)、125.5 (C-6)、124.8 (C-2)、80.8 (C-5)、76.4 (C-4)、19.1 (C-8)。该化合物的比旋光值为
$[\alpha]_{\rm{D}}^{20} $ +78 (c 0.1, MeOH),将该化合物核磁数据与参考文献[17]对照基本一致,确定化合物为terrein。3. 活性测试
对分离得到的化合物进行α-葡萄糖苷酶抑制活性的测试。采用PBS缓冲液为反应体系,利用α-葡萄糖苷酶,以4-硝基苯基-α-D吡喃葡萄糖苷(PNPG)为特异性底物,以阿卡波糖作为阳性药,分别设立空白对照组、α-葡萄糖苷酶空白组和PNPG空白组,评价化合物的α-葡萄糖苷酶的抑制活性。结果表明,化合物3具有较强的α-葡萄糖苷酶的抑制活性,IC50值为14.28 µmol/L。其他化合物没有明显的α-葡萄糖苷酶的抑制活性。另外,还对化合物的抗氧化活性进行测试。采用DPPH的方法,以抗氧化剂N-乙酰半胱氨酸作为阳性药对分离得到的化合物进行了体外抗氧化活性测试。结果显示这些化合物抗氧化活性不明显。
4. 讨论
本研究从棕色扁海绵共附生真菌土曲霉中分离得到了8个化合物,其中化合物3、4、5、7为首次从该菌中分离得到,丰富了土曲霉次级代谢产物的多样性,为进一步探索该属真菌的化学成分和生源途径提供了理论依据。
根据文献报道,化合物2可以提高胰岛素的敏感性[13],化合物4和5测试了多个肿瘤细胞系,均显示细胞毒活性不明显[15],化合物6对大肠杆菌、金黄色葡萄球菌、黄体微球菌、白色念珠菌和隐球菌等具有很好的抗菌活性[16],化合物7对PaCa-2胰腺细胞的抗癌活性和抗菌活性都不明显[17],化合物8能够抑制雄激素依赖性前列腺癌细胞LNCaP-CR的血管生成素分泌,能够抑制人脐静脉内皮细胞的血管形成[18]。为了更好的探究该真菌代谢产物的活性,对分离得到的化合物进行了α-葡萄糖苷酶抑制活性和抗氧化活性测试。其中化合物3显示了较强的α-葡萄糖苷酶的抑制活性,IC50值为14.28 µmol/L,其α-葡萄糖苷酶抑制活性的机制有待于进一步研究。
-
图 2 麻黄碱对PC12细胞形态变化的影响
A. 空白对照;B. 10 μmol麻黄碱;C. 500 μmol麻黄碱浓度;D. 1000 μmol麻黄碱浓度;标尺:500 μm
-
[1] WELLMAN P J, MILLER D K, LIVERMORE C L, et al. Effects of (-)-ephedrine on locomotion, feeding, and nucleus accumbens dopamine in rats[J]. Psychopharmacology (Berl),1998,135(2):133-140. [2] MILLER D K, NATION J R, WELLMAN P J. Sensitization of anorexia and locomotion induced by chronic administration of ephedrine in rats[J]. Life Sci,1999,65(5):501-511. doi: 10.1016/S0024-3205(99)00271-4 [3] SCHMUED L C, BOWYER J F. Methamphetamine exposure can produce neuronal degeneration in mouse hippocampal remnants[J]. Brain Res,1997,759(1):135-140. doi: 10.1016/S0006-8993(97)00173-X [4] BOWYER J F, HOPKINS K J, JAKAB R, et al. L-ephedrine-induced neurodegeneration in the parietal cortex and thalamus of the rat is dependent on hyperthermia and can be altered by the process of in vivo brain microdialysis[J]. Toxicol Lett,2001,125(1-3):151-166. doi: 10.1016/S0378-4274(01)00440-4 [5] LOHOF A M, IP N Y, POO M M. Potentiation of developing neuromuscular synapses by the neurotrophins NT-3 and BDNF[J]. Nature,1993,363(6427):350-353. doi: 10.1038/363350a0 [6] EL-HUSSEINI A E, SCHNELL E, CHETKOVICH D M, et al. PSD-95 involvement in maturation of excitatory synapses[J]. Science,2000,290(5495):1364-1368. doi: 10.1126/science.290.5495.1364 [7] HANSEN S, JØRGENSEN J, NYENGAARD J, et al. Early life vitamin C deficiency does not alter morphology of hippocampal CA1 pyramidal neurons or markers of synaptic plasticity in a Guinea pig model[J]. Nutrients,2018,10(6):749. doi: 10.3390/nu10060749 [8] 郑芳昊, 罗佳波. 麻黄对大鼠中枢神经系统毒副作用的研究[J]. 时珍国医国药, 2015, 26(3):534-536. [9] 郑芳昊, 罗佳波. 麻黄对大鼠额叶皮层氧化损伤的影响[J]. 时珍国医国药, 2016, 27(6):1313-1316. [10] VICARIO-ABEJÓN C, COLLIN C, MCKAY R D, et al. Neurotrophins induce formation of functional excitatory and inhibitory synapses between cultured hippocampal neurons[J]. J Neurosci,1998,18(18):7256-7271. doi: 10.1523/JNEUROSCI.18-18-07256.1998 [11] SHERWOOD N T, LO D C. Long-term enhancement of central synaptic transmission by chronic brain-derived neurotrophic factor treatment[J]. J Neurosci,1999,19(16):7025-7036. doi: 10.1523/JNEUROSCI.19-16-07025.1999 [12] NARISAWA-SAITO M, CARNAHAN J, ARAKI K, et al. Brain-derived neurotrophic factor regulates the expression of AMPA receptor proteins in neocortical neurons[J]. Neuroscience,1999,88(4):1009-1014. doi: 10.1016/S0306-4522(98)00496-5 [13] NARISAWA-SAITO M, SILVA A J, YAMAGUCHI T, et al. Growth factor-mediated Fyn signaling regulates alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor expression in rodent neocortical neurons[J]. Proc Natl Acad Sci USA,1999,96(5):2461-2466. doi: 10.1073/pnas.96.5.2461 [14] CUNHA C, BRAMBILLA R, THOMAS K L. A simple role for BDNF in learning and memory[J]. Front Mol Neurosci,2010(3):1. [15] TAFT C E, TURRIGIANO G G. PSD-95 promotes the stabilization of young synaptic contacts[J]. Philos Trans R Soc Lond B Biol Sci,2014,369(1633):20130134. [16] LIM I A, MERRILL M A, CHEN Y C, et al. Disruption of the NMDA receptor-PSD-95 interaction in hippocampal neurons with no obvious physiological short-term effect[J]. Neuropharmacology,2003,45(6):738-754. doi: 10.1016/S0028-3908(03)00276-4 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4840
- HTML全文浏览量: 1366
- PDF下载量: 21
- 被引次数: 0

