

 下载:
下载:
 下载:
下载:

-
胶原蛋白主要存在于动物皮肤、骨骼和结缔组织中,是主要的结构蛋白质。胶原蛋白具有较低的免疫原性、较好的生物相容性、良好的生物降解性等生物学优点,广泛应用于生物制药、医疗保健、组织工程、食品加工、美容化妆品等领域。目前胶原蛋白主要来源于陆生牲畜(牛、羊和猪等),但某些疾病(口蹄疫、疯牛病)以及宗教相关问题,使得这些牲畜来源的胶原蛋白应用受到一定约束[1]。海洋来源的胶原蛋白资源丰富,且不存在传染性病菌和伦理等问题,因而,对海洋生物来源的胶原蛋白进行开发利用具有良好的应用前景[2]。
目前,从海洋生物提取胶原蛋白的研究主要集中在鳕鱼、罗非鱼、海参以及水母等,这些海洋生物的皮肤、肌肉、软骨组织及中胶层等部位中含有丰富的胶原蛋白[3]。水母是一类胶质状的浮游生物,属刺胞动物门,种类多,分布广,我国近海海域水母资源丰富。大型水母伞部直径可达2 m,中胶层非常厚实,其中胶原蛋白(122.64~693.92 mg/g,干重)约占总蛋白含量的50%[4]。本研究选用青岛海域采集的越前水母(Nemopilema nomurai),对其伞部胶原蛋白进行分离纯化和表征,为水母胶原蛋白的开发应用提供理论基础。
-
SCIENTZ-12N冷冻干燥机(宁波新芝生物科技股份有限公司);LA8080氨基酸自动分析仪(日本株式会社日立高新技术科学);Nicolet iS50傅里叶变换红外光谱仪(美国Thermo Scientific公司);X′Pert3 Powder X射线衍射仪(荷兰帕纳科公司);VARIOSKAN LUX多功能酶标仪(美国Thermo Scientific公司);Mini-PROTEAN Tetra System垂直电泳系统(美国BIO-RAD公司)。
-
越前水母样品采集于青岛海域,腌制后于−20 ℃保存;牛I型胶原蛋白标准品(河北考力森公司);胃蛋白酶(上海生工生物工程公司);标准蛋白Marker(日本TaKaRa公司);其余试剂均为国产分析纯。
-
提取:经腌制的水母样品用自来水清洗,去除表面盐分,4 ℃下用去离子水浸泡、洗净后,再用异丙醇去除脂肪、NaOH去除杂蛋白、过氧化氢去除色素。称取去杂水母伞盖样品,组织匀浆后加入相应体积不同浓度(0.3~0.6 mol/L)的乙酸溶液,胃蛋白酶的添加量为水母伞盖质量的0.1%~0.5%,酶消化时间为24~96 h,反应后的液体使用200目网过滤,离心(4 ℃,10000 r/min)30 min取上清即得粗提液。
纯化:①超滤:分别取粗提液5 ml至分子量10万和5万的超滤管中,4 ℃离心(4000 r/min,30 min),收集滤液进行SDS-PAGE分析。②透析:分别取粗提液5 ml到分子量5万和10万的透析袋中,4 ℃透析3 d,每8 h换一次透析液,取样进行电泳分析。③盐析:取粗提液5 ml,添加NaCl至浓度为0.9 mol/L,4 ℃下10000 r/min离心10 min,收集沉淀,复溶于0.5 mol/L乙酸溶液,重复上述过程2次,取溶液进行电泳分析。仅改变NaCl浓度(0.9 mol/L→0.9 mol/L→2.0 mol/L或2.0 mol/L→0.9 mol/L→0.9 mol/L),重复实验。
粗提液纯化后,在−80 ℃冷冻干燥得水母酶溶性胶原蛋白(PSC)。
-
以水母胶原蛋白得率(冻干胶原蛋白质量/水母干重)为指标,确定基本反应条件(乙酸浓度、料液比、胃蛋白酶用量、提取时间),每次改变其中一个条件进行水母胶原蛋白提取的单因素实验。
获得单因素数据后,设计正交实验(表1)来获取最佳的实验方案。
表 1 水母胶原蛋白提取正交实验因素水平表
水平 因素 A加酶量(%) B乙酸浓度(mol/L) C料液比 D反应时间(h) 1 0.1 0.3 1∶1 24 2 0.2 0.4 1∶2 48 3 0.3 0.5 1∶3 72 -
参照Coentro等[5]的方法检测胶原蛋白浓度,将100 μl胶原蛋白溶液添加至1 ml天狼星红色染料中静置30 min,然后以12000 r/min离心30 min。去除上清,沉淀用碱性试剂溶解30 min,取200 μl胶原蛋白液于540 nm处读数。
-
用X射线衍射仪分析胶原蛋白样品的晶体结构,X射线源为Cu靶Ka辐射(λ=0.154 nm),电流为40 mA,电压为40 kV。扫描角度2θ为5°~40°,扫描速率为3.6°/min,并根据公式λ=2d sinθ,计算d值。
-
胶原蛋白溶解于0.5 mol/L乙酸中,用多功能酶标仪检测其紫外光谱。扫描波段为200~400 nm,波长间隔为1 nm。以0.5 mol/L乙酸溶液作为空白对照。
-
取胶原蛋白5 mg和适量KBr置于研钵中,研磨均匀。取适量的混合样品于压片磨具中,用压片机制成试样薄片。将压片放入傅里叶变换红外光谱仪内进行扫描(500~4000 cm−1,分辨率2 cm−1)。
-
胶原蛋白溶液和上样缓冲液混匀后水浴(100 ℃)加热5 min,取15 μl上样,使用8%的分离胶和5%的浓缩胶,80 V电泳100 min。
-
胶原蛋白样品在110 ℃下盐酸水解24 h,水解产物12000 r/min离心20 min,上清液用氨基酸自动分析仪进行氨基酸含量测定。
-
将胶原蛋白样品溶于0.5 mol/L乙酸溶液中,配成1 mg/ml的胶原蛋白溶液,分别取10 ml溶液调配成不同pH值(3~9共7个梯度)和NaCl浓度(0.5、0.6、0.7、0.8、0.9、1.0、1.1 mol/L),10000 r/min离心,取上清液进行胶原蛋白浓度测定,并绘制变化曲线。
-
数据采用(
$\bar x $ ±s)表示,用 GraphPad Prism 8.0.2作图并对各组进行方差分析,两组间比较采用t检验。检验水准α=0.05,P<0.05有统计学意义。 -
从图1A可见,当酶的用量从0.1%升到0.3%时,胶原蛋白得率逐渐增高并达到最大值。继续增加酶使用量,胶原蛋白得率反而减少,可能是过量酶对胶原蛋白具有一定破坏作用。从图1B可见,当乙酸浓度为0.4 mol/L时,胶原蛋白的得率最高。而乙酸过量,会破坏已生成胶原蛋白。从图1C可见,料液比在1∶2 g/ml时,水母胶原蛋白的得率最大。可能是料液比低于1∶2 g/ml时,胶原溶胀不充分,而料液比继续增加并没有多余胶原以供溶胀。从图1D可见,随着时间推移,胶原蛋白的得率先升高再下降:48 h和24 h相比,增加了8.8%;72 h和48 h相比,增加了15.9%;96 h和72 h相比,得率反而下降,说明72 h是胶原蛋白提取的最佳时间点。
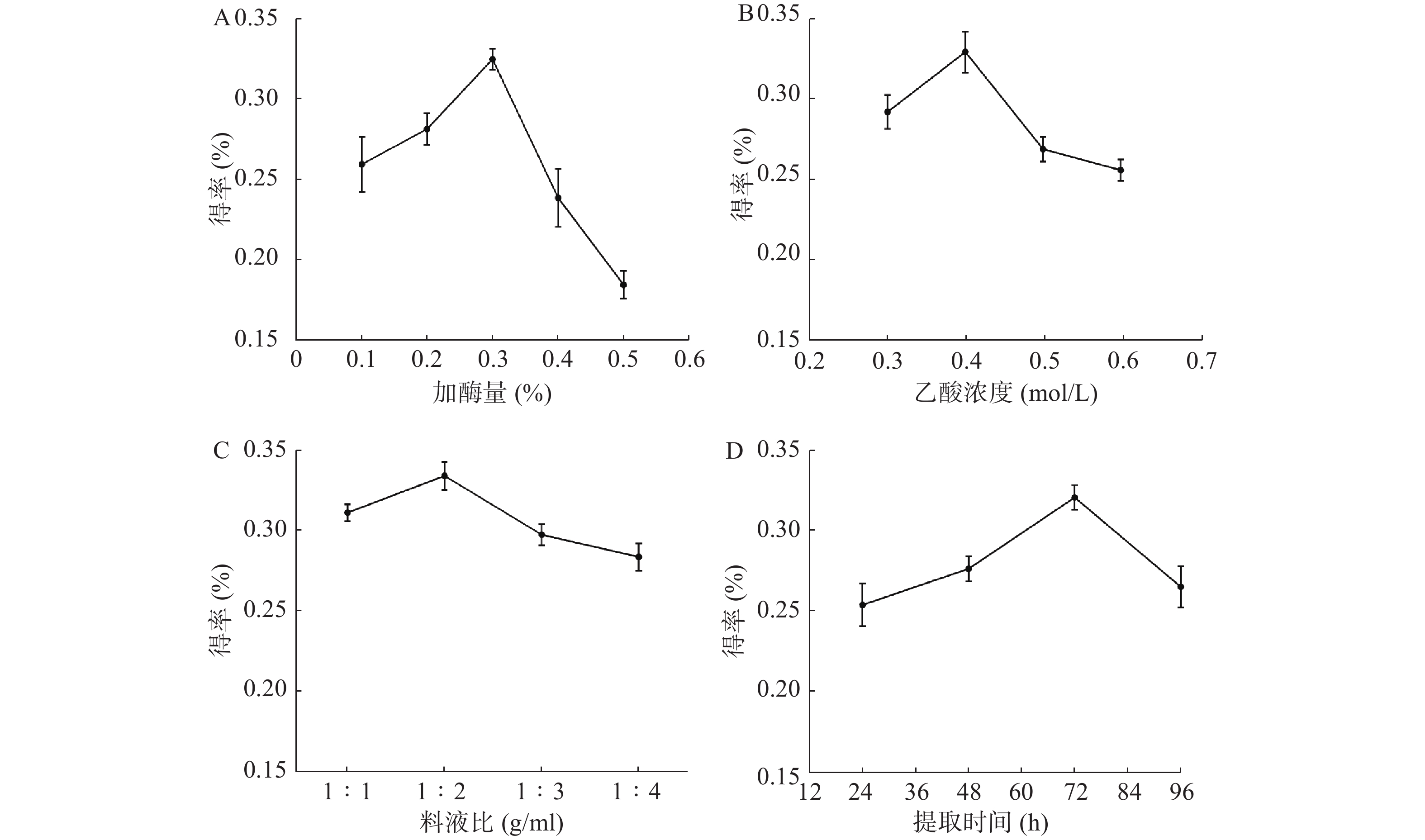
图 1 水母胶原蛋白提取的单因素分析
-
表2正交实验极差R分析中,RA>RD>RC>RB,即在胶原蛋白提取中,酶的使用量起着最关键的作用,提取时间排第二,料液比和乙酸浓度的影响比较小。综合这9组数据,确定最优理论水平为A3B2C2D3,即乙酸(0.4 mol/L)、料液比(1∶2 g/ml)、胃蛋白酶(0.3%),提取72 h,此时的得率为(33±0.63)%。
表 2 水母胶原蛋白提取正交实验结果
水平 因素 A 加酶量(%) B 乙酸浓度(mol/L) C料液比(g/ml) D时间(h) 胶原蛋白得率(%) 实验1 0.1 0.3 1∶1 24 17.8±0.66 实验2 0.1 0.4 1∶2 48 23.2±0.94 实验3 0.1 0.5 1∶3 72 21.7±0.85 实验4 0.2 0.3 1∶2 72 26.7±0.58 实验5 0.2 0.4 1∶3 24 19.0±0.86 实验6 0.2 0.5 1∶1 48 24.2±0.76 实验7 0.3 0.3 1∶3 48 26.9±0.61 实验8 0.3 0.4 1∶1 72 31.1±0.50 实验9 0.3 0.5 1∶2 24 24.3±0.75 K1 20.9 23.8 24.3 20.4 K2 23.3 24.4 24.7 24.8 K3 27.4 23.4 22.5 26.5 R 6.53 1.03 2.20 6.13 因素主次 A>D>C>B 最佳组合 A3B2C2D3 -
图2中水母胶原蛋白粗提物(泳道8)在分子量13万处含有大量的目标胶原蛋白,分子量10万以下有杂条带。不同规格透析袋处理(泳道1、2)后,分子量10万以下的杂蛋白部分去除;盐析结果显示,0.9 mol/L→0.9 mol/L→2.0 mol/L NaCl的盐析(泳道4)可以去除大量杂蛋白,但目标蛋白损失较多,而0.9 mol/L→0.9 mol/L→0.9 mol/L NaCl(泳道3)处理既可以保留目标蛋白又能够去除部分杂蛋白。最后超滤结果显示,分子量10万(泳道6)的超滤管可以明显减少杂蛋白含量。
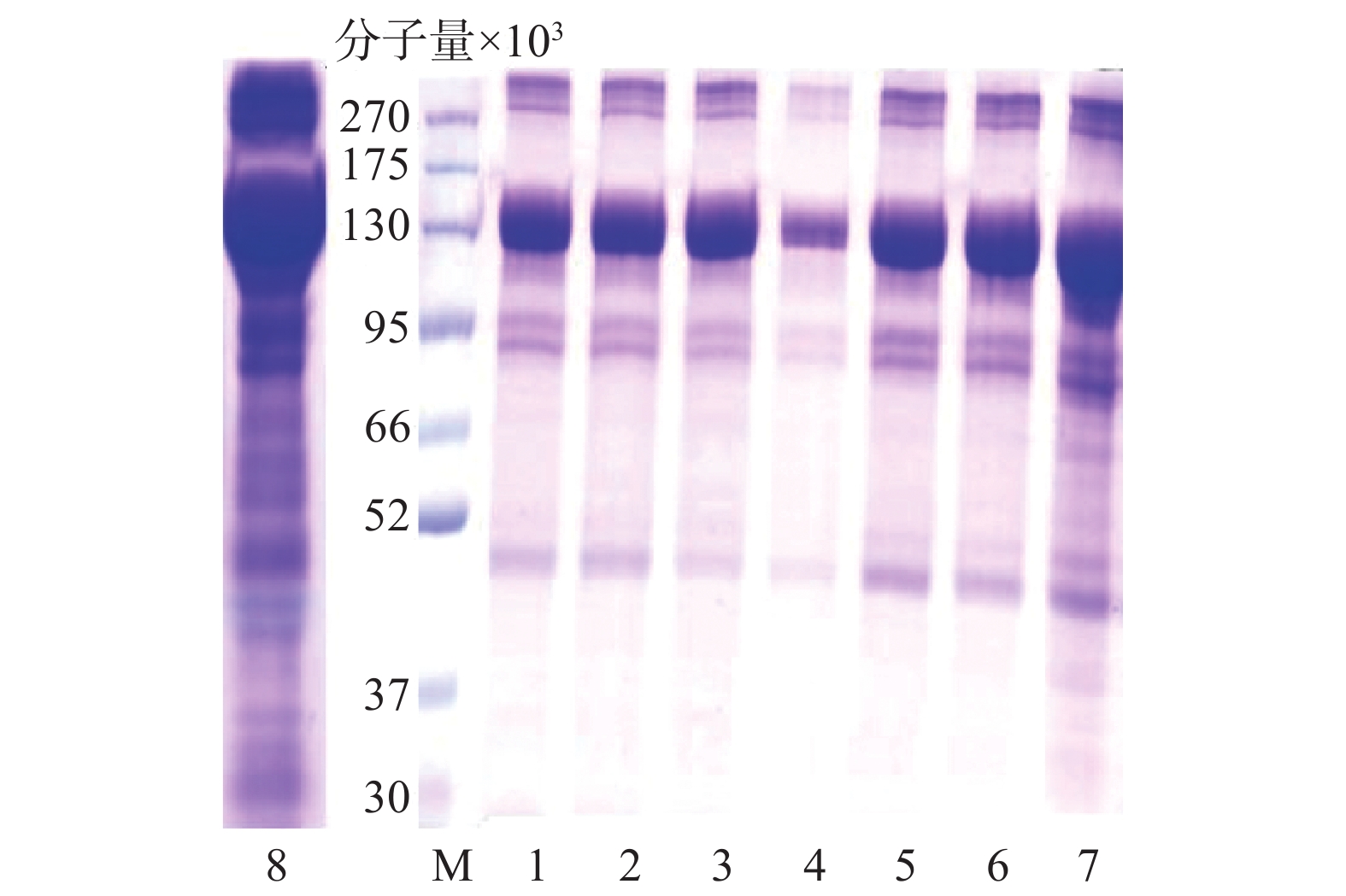
图 2 水母胶原蛋白纯化过程SDS-PAGE电泳图
-
纯化实验结果提示,单一纯化方法不能完全去除杂蛋白,传统透析处理耗时长,而反复盐析会增加胶原蛋白溶液NaCl含量。综合权衡后确定纯化条件为0.9 mol/L→0.9 mol/L→0.9 mol/L NaCl盐析和100 kDa超滤组合。图3是组合纯化后胶原蛋白电泳图:泳道b为牛I型胶原蛋白标准品,在分子量13万左右有α1、α2链,而β链γ链的分子量都在20万以上。本实验提取的水母胶原蛋白(泳道a),分子量10万下基本没有杂蛋白,但α链只发现了一条,其分子可能是(α1)3[6-7],分子量在13万左右,符合I型胶原蛋白的特征。
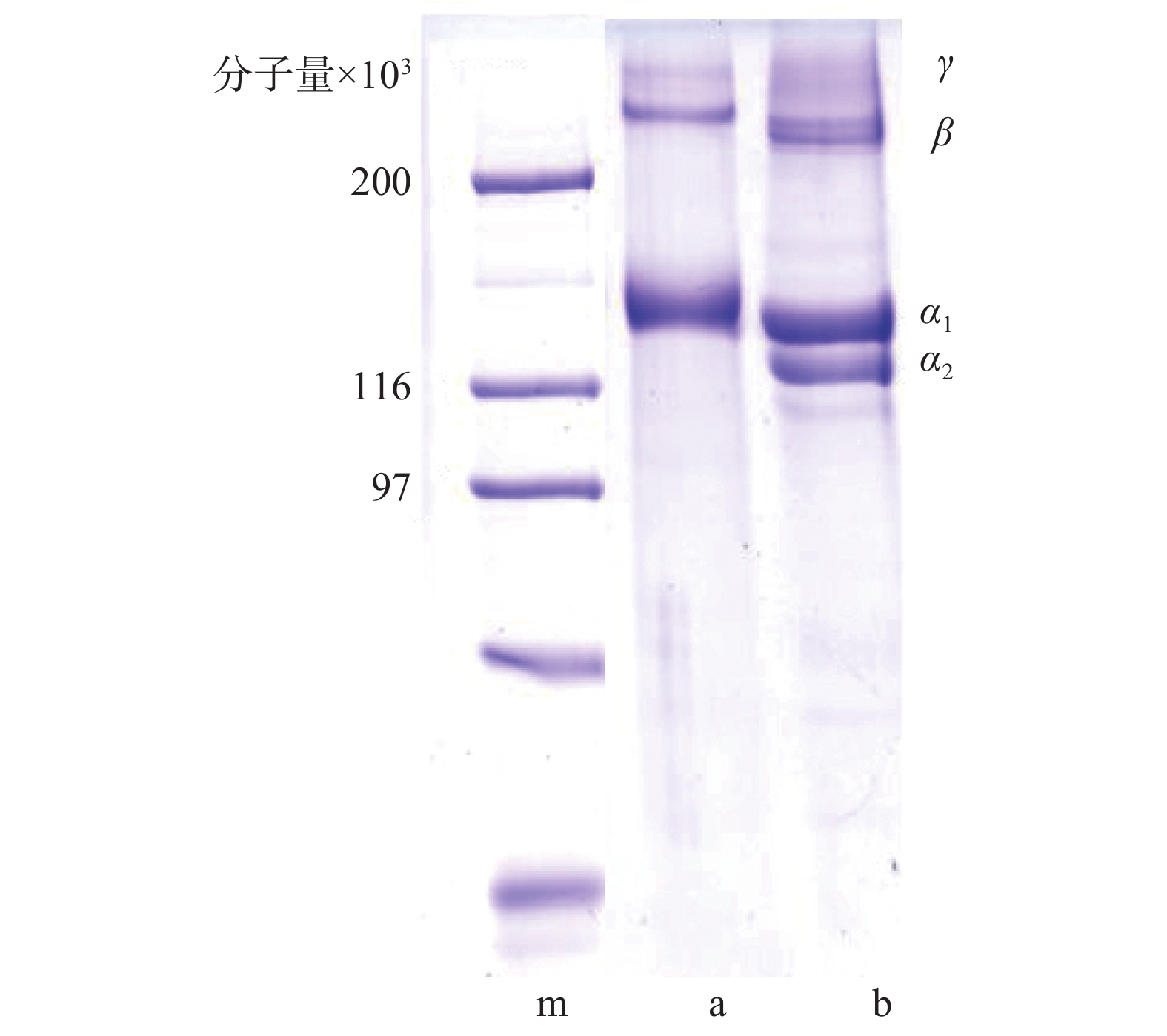
图 3 纯化后水母胶原蛋白SDS-PAGE电泳图
-
胶原蛋白X射线衍射分析结果如图4、表3所示。水母胶原蛋白和牛胶原蛋白都有3个衍射峰。峰1形状尖锐与胶原蛋白三螺旋结构有关,d值反映胶原蛋白分子链间距离[8],水母和牛胶原蛋白的d1值分别为1.244 nm和1.226 nm。在20°附近出现一个宽大峰(峰2),代表胶原蛋白纤维内部众多结构层次所引起的漫散射[9]。在30°附近出现了第3个峰(峰3),该峰较小,其d值显示三股螺旋结构中沿螺旋中心轴相邻氨基酸残基之间的距离[10],水母和牛胶原蛋白峰3的d3值分别为0.295 nm和0.296 nm,极为接近。说明水母胶原蛋白和牛胶原蛋白标准品一样,保持了完整的三股螺旋结构。
表 3 胶原蛋白X射线衍射峰对应的d值
组别 衍射峰 2θ/(°) d值(nm) 水母胶原蛋白 峰1 7.1 1.244 峰2 21.3 0.418 峰3 30.2 0.295 牛胶原蛋白 峰1 7.2 1.226 峰2 20.3 0.437 峰3 30.1 0.296 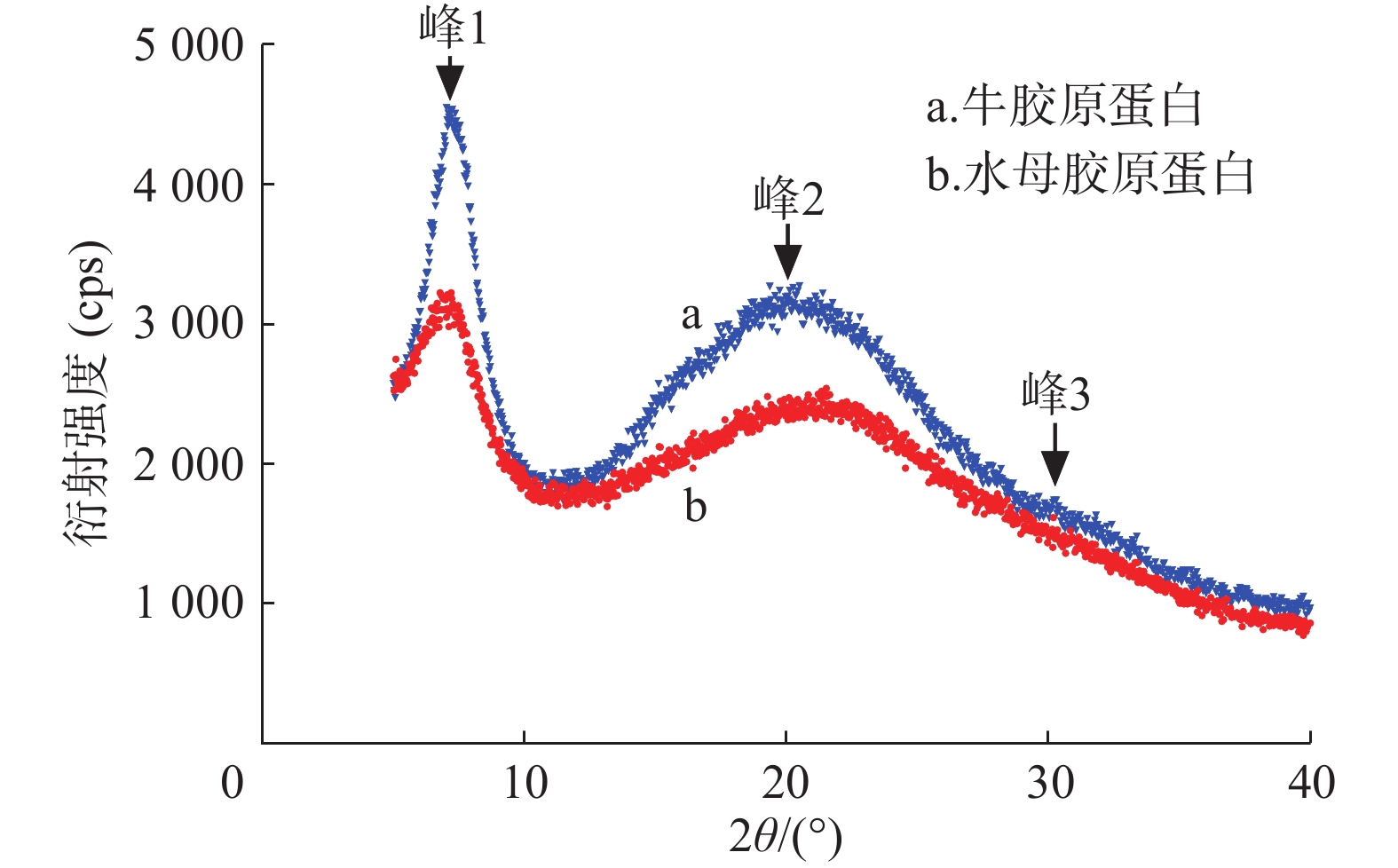
图 4 胶原蛋白的X射线衍射图谱
-
蛋白通常含有芳香族氨基酸酪氨酸、色氨酸和苯丙氨酸,这些氨基酸含有苯环共轭双键系统,在特定波长(280 nm和251 nm)处有最大吸收峰。但在胶原蛋白中这些氨基酸含量少,其紫外吸收峰主要集中在220~230 nm,主要是由羰基C=O的n→π跃迁所致。水母胶原蛋白的紫外光谱如图5所示,其最强吸收峰在220 nm处,与牛胶原蛋白吸收峰出现的位置类似,接近Ab Aziz等[11]报道的水母Rhopilema hispidum胶原蛋白紫外吸收峰(218 nm),提示本实验中提取的水母胶原蛋白纯度较高。
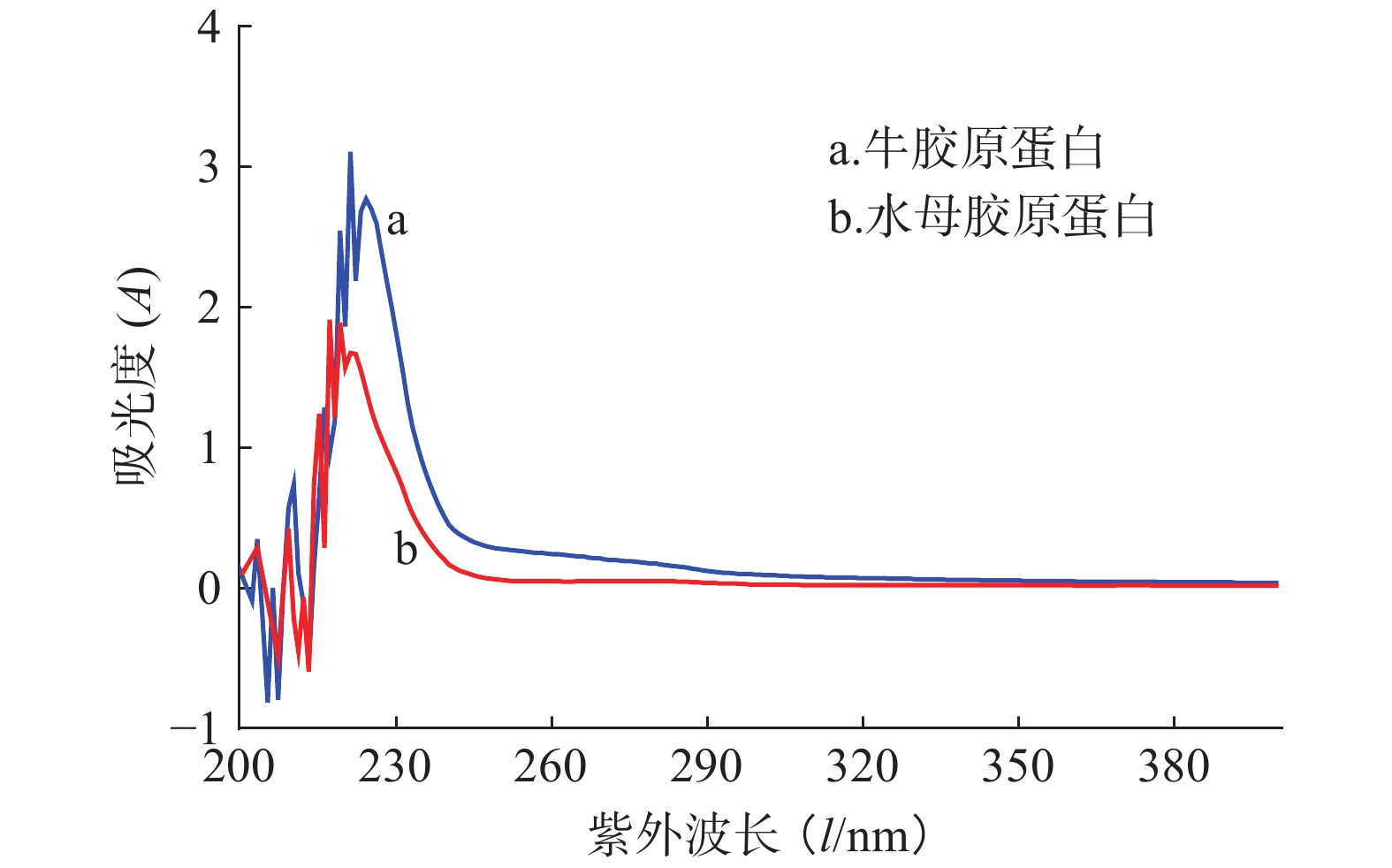
图 5 胶原蛋白紫外光谱
-
从图6可见,水母胶原蛋白酰胺A吸收峰为3288 cm−1,和Rastian等[12]检测的酰胺A(3292 cm−1)吸收峰接近,表明氢键存在于胶原蛋白结构中。胶原蛋白分子中CH2基团的不对称伸缩振动产生了酰胺B特征吸收峰[13-14],水母胶原蛋白的酰胺B在2933 cm−1处,和牛胶原蛋白的酰胺B(2936 cm−1)吸收峰类似。作为蛋白质二级结构变化的特征区域——酰胺Ⅰ带(1625~1690 cm−1),是由蛋白质肽链骨架的C=O伸缩振动产生[15],水母胶原蛋白的酰胺I带出现在1641 cm−1。水母胶原蛋白酰胺II带吸收峰出现在1543 cm−1,在胶原蛋白酰胺II带(1500~1600 cm−1)的范围内,可能是C—N键的伸缩振动和N—H弯曲振动产生。水母胶原蛋白酰胺Ш位于1236 cm−1处,主要是脯氨酸侧链和甘氨酸骨架的—CH2摇摆振动产生,提示所提取的水母胶原蛋白三螺旋结构完整性较好。
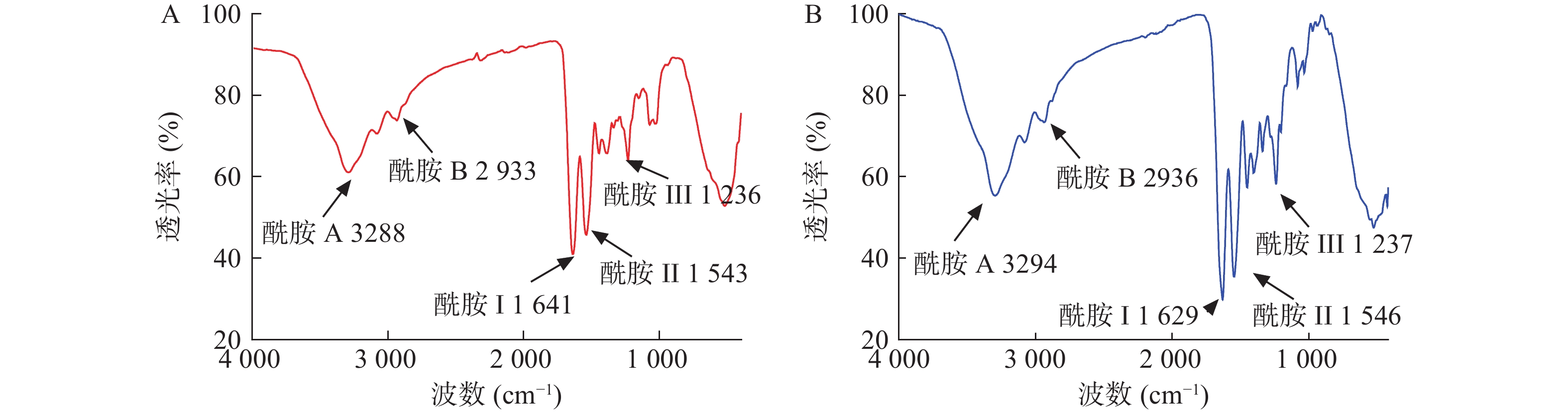
图 6 胶原蛋白的红外光谱
-
电泳图谱表明提取的水母胶原蛋白达到电泳纯(95%)。文献报道[16-18]天狼星红染料与胶原蛋白中的特征性结构Gly-X-Y结合,可以使用天狼星红染料对胶原蛋白进行快速检测。本实验测得每100 mg冻干的水母胶原蛋白样品中含有胶原蛋白96.85 mg,其纯度为96.85%。
-
如表4所示,水母胶原蛋白的氨基酸组成中,甘氨酸占34.8%,与云南深水鲷鱼皮胶原蛋白甘氨酸含量(35%)接近,比热带淡水鲤鱼鳞中胶原蛋白甘氨酸含量(30.6%)高[19]。水母的胶原蛋白中羟脯氨酸占6.3%,脯氨酸占8.2%,亚氨酸比例为14.5%。其脯氨酸羟基化度(羟脯氨酸含量/亚氨酸含量)为43.68%,比牛胶原蛋白标准品的羟基化度(39.92%)和Zhang等[7]提取的水母胶原蛋白的羟基化度(32.2%)都高。据文献报道[20],羟基化度越高,胶原蛋白的三螺旋结构越稳定。检测结果中还发现蛋氨酸、酪氨酸和组氨酸的含量比较低,缺乏色氨酸,这些都符合I型胶原蛋白的特点[21]。
表 4 二种胶原蛋白中不同氨基酸组分的百分比
氨基酸名称 氨基酸占比(%) 水母胶原蛋白 牛胶原蛋白 天冬氨酸Asp 7.83 4.57 苏氨酸Thr 3.55 1.73 丝氨酸Ser 4.89 3.19 谷氨酸Glu 9.98 7.27 甘氨酸Gly 34.82 34.37 丙氨酸Ala 8.36 12.46 缬氨酸Val 2.74 2.27 蛋氨酸Met 0.21 0.01 异亮氨酸Ile 1.88 1.22 亮氨酸Leu 3.09 2.64 酪氨酸Tyr 0.11 0.27 苯丙氨酸Phe 0.9 1.38 赖氨酸Lys 2.96 2.15 组氨酸His 0.29 0.48 精氨酸Arg 3.87 5.3 脯氨酸Pro 8.16 12.43 羟脯氨酸Hyp 6.33 8.26 合计 100 100 注:氨基酸分析采用氨基酸残基数占100个氨基酸残基的比例 -
从图7A可见,当NaCl浓度在0.5~0.8 mol/L时,水母胶原蛋白相对溶解度比较稳定,保持着最大值。其原因可能是低浓度的盐离子可以与胶原蛋白分子结合,使得胶原蛋白分子带有更多的电荷,相互排斥,溶解度较大。当NaCl浓度达到0.9 mol/L时,水母胶原蛋白的溶解度急剧下降,相对溶解度不到23%,当NaCl浓度超过1.0 mol/L后,其相对溶解度在15%左右。可能因为NaCl的浓度较高时,极性较强的盐离子夺走了胶原蛋白结合的水分子,破坏了其周围的水化膜,盐析作用显现出来,溶解度下降。
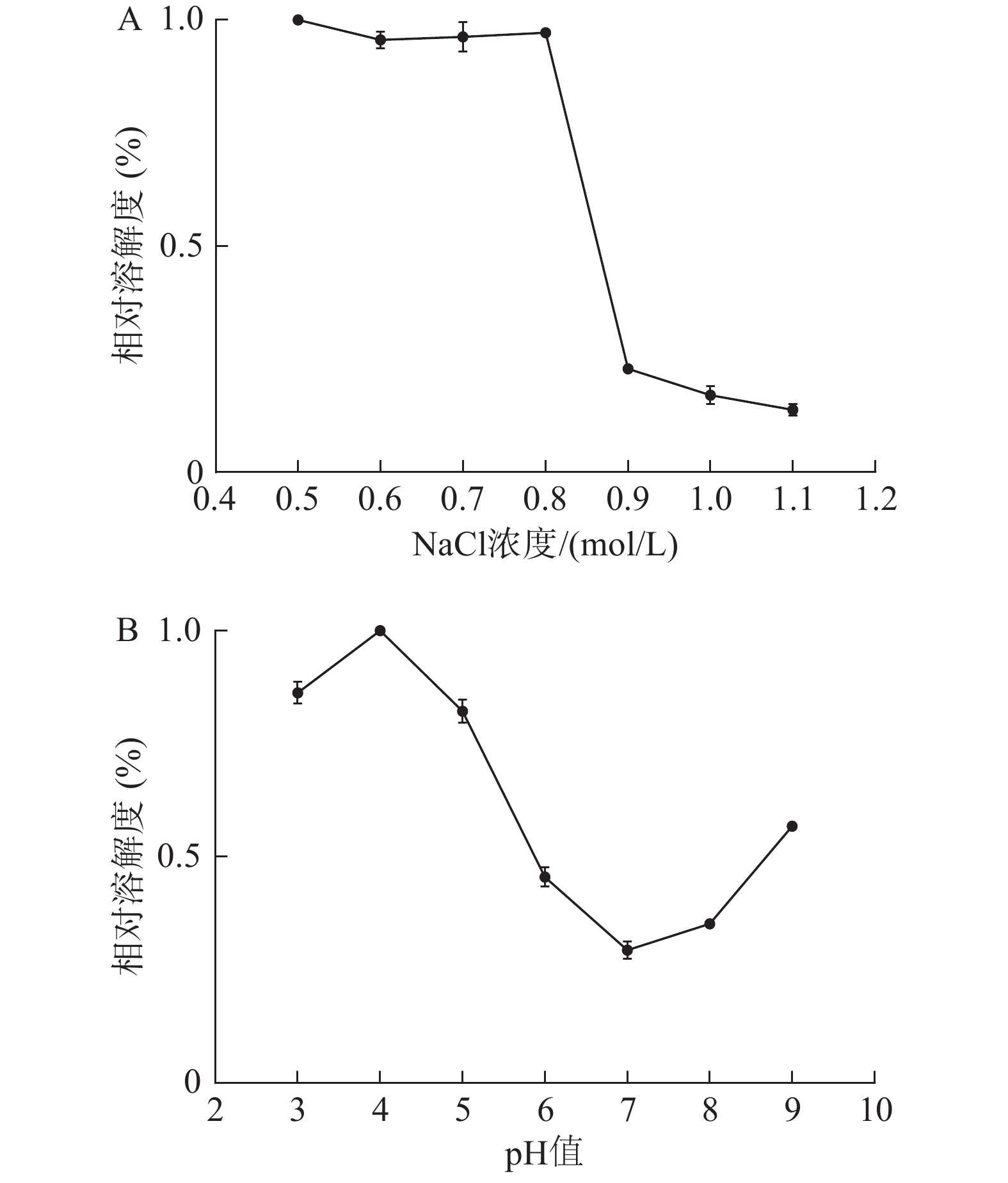
图 7 水母胶原蛋白在不同pH值及NaCl浓度条件下溶解度特性
从图7B可见,当溶液的pH在3~5时,水母胶原蛋白的相对溶解度比较大,维持在80%左右。其中pH为4时,溶解度最大,与海参胶原蛋白的溶解性类似[3]。当pH在6~8时,胶原蛋白的相对溶解度维持在较低值,尤以pH 7时溶解度最小,此时就是胶原蛋白的等电点。当pH超过8后,胶原蛋白溶液的溶解度又有所上升。
-
本实验通过单因素分析和正交实验优化了水母胶原蛋白的提取条件,确定最适条件为:乙酸0.4 mol/L、料液比1∶2 g/mL、胃蛋白酶0.3%、反应72 h。水母胶原蛋白的纯化方法优化为0.9 mol/L-0.9 mol/L-0.9 mol/L NaCl盐析和分子量10万超滤的组合方法。相比阳离子交换层析法和Sephacryl S-100凝胶过滤纯化方法,超滤不仅可以去除盐分和杂蛋白,还能快速富集胶原蛋白,缩短提取时间。
纯化的水母胶原蛋白通过天狼星红法检测其纯度为97%,高于从其他多种海洋生物提取的I型胶原蛋白纯度[22-23]。本实验中越前水母胶原蛋白干重得率为33%左右,相比于水母Chrysaora quinquecirrha胶原蛋白提取得率(1.2%干重)显著提高[24]。凝胶电泳显示提取的水母胶原蛋白构型可能是(α1)3,α1链分子量约为13万和10万以下基本没有杂条带,比Jankangram等[25]提取的水母胶原蛋白纯度更高。在氨基酸组成分析中,越前水母胶原蛋白甘氨酸占34.82%,羟脯氨酸占6.33%,脯氨酸占8.16%,没有检测到色氨酸。以上结果均表明水母胶原蛋白符合I型胶原蛋白的特点。光谱分析发现,分离纯化过程中水母胶原蛋白三螺旋结构保存较好。溶解度试验表明其在pH=4时溶解性最佳,当NaCl浓度升高至0.9 mol/L时溶解度快速下降。
总之,本研究通过优化提取纯化过程,缩短了水母胶原蛋白制备时间,提高了所得胶原蛋白的纯度,且水母胶原蛋白三螺旋结构保持完整,与牛I型胶原蛋白特征相似度很高,提示水母胶原蛋白可望成为哺乳动物胶原蛋白的替代品。鉴于水母胶原蛋白具有资源丰富、提取方便、生物相容性好等优点,可望成为生物医药、食品、化妆品等多领域的理想生物材料,具有良好的应用前景。
Isolation and characterization of collagen from the jellyfish Nemopilema nomurai
-
摘要:
目的 优化越前水母(Nemopilema nomurai)胶原蛋白提取工艺,并探究其性能。 方法 利用乙酸和胃蛋白酶提取胶原蛋白,通过盐析和超滤纯化水母胶原蛋白粗提物,并对水母胶原蛋白进行表征和性能分析。 结果 水母胶原蛋白纯度97%,得率33%(干重);X射线衍射、紫外光谱和傅里叶变换红外光谱检测结果表明,水母胶原蛋白在提取和纯化过程中保持了胶原蛋白三螺旋结构;氨基酸组成和凝胶电泳分析提示,水母胶原蛋白符合Ⅰ型胶原蛋白特征。溶解度实验表明,水母胶原蛋白在pH 3~5,NaCl浓度小于0.9 mol/L的条件下都具有良好的溶解性。 结论 所得水母胶原蛋白与牛Ⅰ型胶原蛋白特征相似,有望成为新的胶原蛋白资源。 Abstract:Objective To optimize the extraction process of collagen from the jellyfish (Nemopilema nomurai) and explore its characters. Methods The collagen was extracted with acetic acid and pepsin from jellyfish. The crude jellyfish collagen was purified by salting out and ultrafiltration. The purified collagen was characterized and analyzed by XRD (X-ray diffraction), UV (ultraviolet spectroscopy) and FTIR (fourier transform infrared spectroscopy). Results The purity of jellyfish collagen was 97%, the yield was 33% (dry weight). The jellyfish collagen maintained its triple helix structure during the extraction and purification process. Jellyfish collagen conformed to the characteristics of type I collagen according to amino acid composition and gel electrophoresis analysis. The jellyfish collagen exhibited a good solubility under the conditions of pH 3–5 and less than 0.9 mol/L of NaCl. Conclusion The extracted jellyfish collagen exhibited similar characteristics with bovine type I collagen and was a potential new source of collagen. -
Key words:
- Nemopilema nomurai /
- jellyfish /
- collagen /
- extraction /
- purification
-
补体系统是人体重要的免疫防御系统之一,是由30多种广泛存在于血清、组织液和细胞膜表面的蛋白质组成的,具有精密调控机制的蛋白质反应系统,其主要通过3种途径激活:经典途径、旁路途径和甘露糖结合凝集素途径。补体系统正常激活,可在靶细胞上形成膜攻击复合物,导致靶细胞的溶解,补体的这一功能在机体的免疫系统中起重要的防御和免疫监视作用,对抵御外来微生物的入侵和维持机体平衡有重要的作用。然而该系统的过度激活将释放炎性过敏毒素C3a和C5a,具有化学诱导作用的C5a能趋化嗜中性粒细胞、中核细胞和嗜酸性粒细胞,这些细胞释放蛋白酶和具有趋化作用细胞因子,进一步聚集T、B淋巴细胞和其他炎性细胞,从而促进炎症反应的发生,引起系统性红斑狼疮、类风湿性关节炎、动脉粥样硬化、肾小球肾炎等[1-2]。近年来已有研究表明[3],补体系统的激活是类风湿性关节炎中慢性滑膜炎的发病因素之一。因此,抑制补体系统的过度激活可能是治疗类风湿性关节炎的重要机制之一。
三色片为复旦大学附属中山医院的院内制剂,由雷公藤、黄芪和丹参三味药材按1∶1∶1的比例配伍组成,在临床上用于治疗类风湿性关节炎、系统性红斑狼疮、银屑病和湿疹等结缔组织疾病。我院临床医生在长期的医疗实践中总结出来的经验方,效果显著[4]。组方中雷公藤,性味辛寒,有大毒,归肝、肾经,具有清热解毒、活血化瘀、通络止痛、杀虫止痒等功效。现代研究表明,雷公藤内酯醇对大鼠脑皮质内注射β-淀粉酶后补体C1q和C3的表达有抑制作用,表明雷公藤对补体系统有抑制作用,目前临床上广泛用于治疗类风湿性关节炎、系统性红斑狼疮、银屑病和湿疹等结缔组织疾病[5]。组方中的黄芪用于脾肺气血或中气下陷之症、卫气虚所致表虚自汗、气虚血滞导致的肢体麻木、关节痹痛等症,可联合治疗类风湿性关节炎[6]。黄芪在治疗2型糖尿病大鼠的研究中发现其能降低补体C3的水平,表明其对补体系统具有一定的调节作用[7-8]。丹参是最常用的活血化瘀中药之一,具有祛瘀止痛,养血安神的功效,现代药理学研究表明其还具有保护肝脏的功能[9],可拮抗雷公藤的肝毒性。本研究通过经典途径抗补体活性测定方法筛选出三色片醇提物的乙酸乙酯部位抗补体活性最佳,并采用UPLC-Q-TOF-MS法对该部位的化学成分进行结构表征,为三色片抗补体活性药效物质基础及治疗补体过度激活相关疾病提供科学依据。
1. 仪器、试剂与材料
Tripie TOF5600+型四级杆-飞行时间串联质谱仪,配备电喷雾电离源和CDS自动校正系统(美国Applied Biosystems公司);Peak view2.2和Master view1.1数据处理系统(美国Applied Biosystems公司);LC-30A超高效液相色谱仪,包括高压输液泵,自动进样器,柱温箱和在线脱气机(日本岛津公司);KQ5200E型超声清洗器(昆山市超声仪器有限公司);甲醇、乙腈(色谱纯,德国Merck公司);甲酸(色谱纯,美国Sigma-Aldrich公司); 蒸馏水(娃哈哈集团);三色片提取物由作者自制,现样品存放于复旦大学附属中山医院药剂科(SSP2018);补体、溶血素(自制);毛蕊异黄酮(批号:ST088101),雷公藤甲素(批号:ST020501),雷公藤内酯酮(批号:ST049901),丹参酮II A(ST014601)、黄芪甲苷(ST001601)(纯度≥ 98%,均购自上海斯丹德生物技术有限公司)。
2. 方法
2.1 三色片醇提物及各极性部位的制备
雷公藤、黄芪和丹参三味药材按1∶1∶1配伍,其中,黄芪和丹参加6倍量的水浸泡2 h后,煎煮2次,第一次1.5 h,第二次加水4倍量煎煮1 h,煎液滤过,合并滤液并浓缩至相对密度为1.10~1.20(70 ℃),加入2倍量的乙醇,静置沉淀24 h,取上清液备用。雷公藤分别加4倍量的乙醇加热回流2次,每次1.5 h,合并提取液,滤过,加入上述备用药液,混匀,回收乙醇至无醇味,浓缩后即得三色片醇提物,经现有的质量标准检验为制备三色片制剂合格的提取物。精密称取三色片醇提物2.0 g,置于100 ml萃取瓶中,加25 ml蒸馏水溶解后,用等量的石油醚、乙酸乙酯和正丁醇进行萃取,浓缩干燥后,放冷至室温,得到三色片醇提物的石油醚部位0.36 g,乙酸乙酯部位0.42 g,正丁醇部位0.56 g和水溶性部位。
2.2 经典途径的抗补体活性测定
取各极性部位样品2 mg溶于DMSO,采用BBS缓冲液稀释成不同浓度的样品,并加入临界浓度的补体(1∶80稀释的豚鼠血清),溶血素和2%绵羊红细胞(SRBC)。37 ℃水浴30 min,离心后取上清液在405 nm波长下测定吸光度(A)值。同时设置中药对照组(将等量的中药提取物加入BBS缓冲液中,用于测定中药本底A值)、补体组(取临界浓度的补体直接加入适量的BBS缓冲液、溶血素和2%SRBC,用于测定临界浓度补体所造成红细胞溶血的A值)和全溶血组(将2%SRBC加入水中使之全溶血,用于观察补体组是否达到或接近全溶血水平),并以肝素作为阳性对照组,计算溶血抑制率。以供试品浓度为横坐标(X),溶血抑制率为纵坐标(Y),计算CH50(经典途径50%抑制溶血所需供试品浓度)。溶血抑制率=1−(A中药−A中药对照)/A全溶血。
2.3 不同浓度的样品色谱与质谱条件
2.3.1 色谱条件
色谱柱为ACQUITY UPLC BEH C18(2.1 mm×100 mm,1.7 μm);流动相0.1%甲酸和水溶液(A)−乙腈(B);梯度洗脱:0~9 min,10%~23% B;9~13 min,23% B;13~28 min,23%~40% B;28~32 min,40%~50% B;32~37 min,50%~100% B;37~42 min,100% B;42~42.1 min,10%B;42.1~50 min,10% B;流速为0.25 ml/min,柱温为35 ℃;进样量为2 μl。
2.3.2 质谱条件
在正/负离子模式,离子源选择电喷雾离子化源(ESI);使用m/z 50~1250扫描范围;碰撞能量35 eV,碰撞能量叠加(35±15)eV;喷雾电压5 500 V;雾化气温度550 ℃;去簇电压100 V;雾化气和辅助气均为50 psi;气帘气25 psi;数据采集时间50 min;采用母离子触发的子离子(TOF-MS-IDA-MS/MS)扫描方式;多重质量亏损和动态背景扣除为触发二级的条件,满足该条件进行二级扫描。
2.4 对照品溶液的制备
精密称取毛蕊异黄酮、雷公藤甲素、雷公藤内酯酮、丹参酮Ⅱ A和黄芪甲苷对照品1.0 mg,加甲醇2 ml,溶解,摇匀,即得各对照品溶液。
2.5 供试品溶液的制备
取三色片醇提物的乙酸乙酯部位样品0.2 g,置于10 ml量瓶中,加入70%甲醇5 ml,超声处理(功率250 W,频率40 kHz)30 min,放冷至室温,70%甲醇定容至刻度,摇匀,滤过,取续滤液,即得供试品溶液。
2.6 三色片中化学成分数据库的建立
根据三色片中各药材化学成分研究文献,收集3种药材所含化合物成分的基本信息,包括化合物名称、分子式、精确分子量、准分子离子峰和碎片离子峰。通过精确分子量匹配,对照品的保留时间,二级谱所得到的离子碎片与文献报道进行比对,最终确定化合物的结构。
3. 结果与分析
3.1 三色片醇提物各极性部位的抗补体活性
分别对三色片醇提物的石油醚部位、乙酸乙酯部位和正丁醇部位进行经典途径的抗补体活性测定,以肝素为对照品,结果发现乙酸乙酯部位的抗补体活性最好,其抗补体活性略低于肝素钠,其次是正丁醇部位,结果见表1。
表 1 三色片提取物不同部位抗补体活性测定编号 研究对象 抗补体活性(CH50,μg/ml) 1 肝素 14.4±1.2 2 三色片-石油醚部位 − 3 三色片-乙酸乙酯部位 233.9±10.1 4 三色片-正丁醇部位 344.0±14.5 注:“—”表示该部位无抗补体活性。 3.2 三色片醇提物乙酸乙酯部位的UPLC-Q-TOF-MS分析
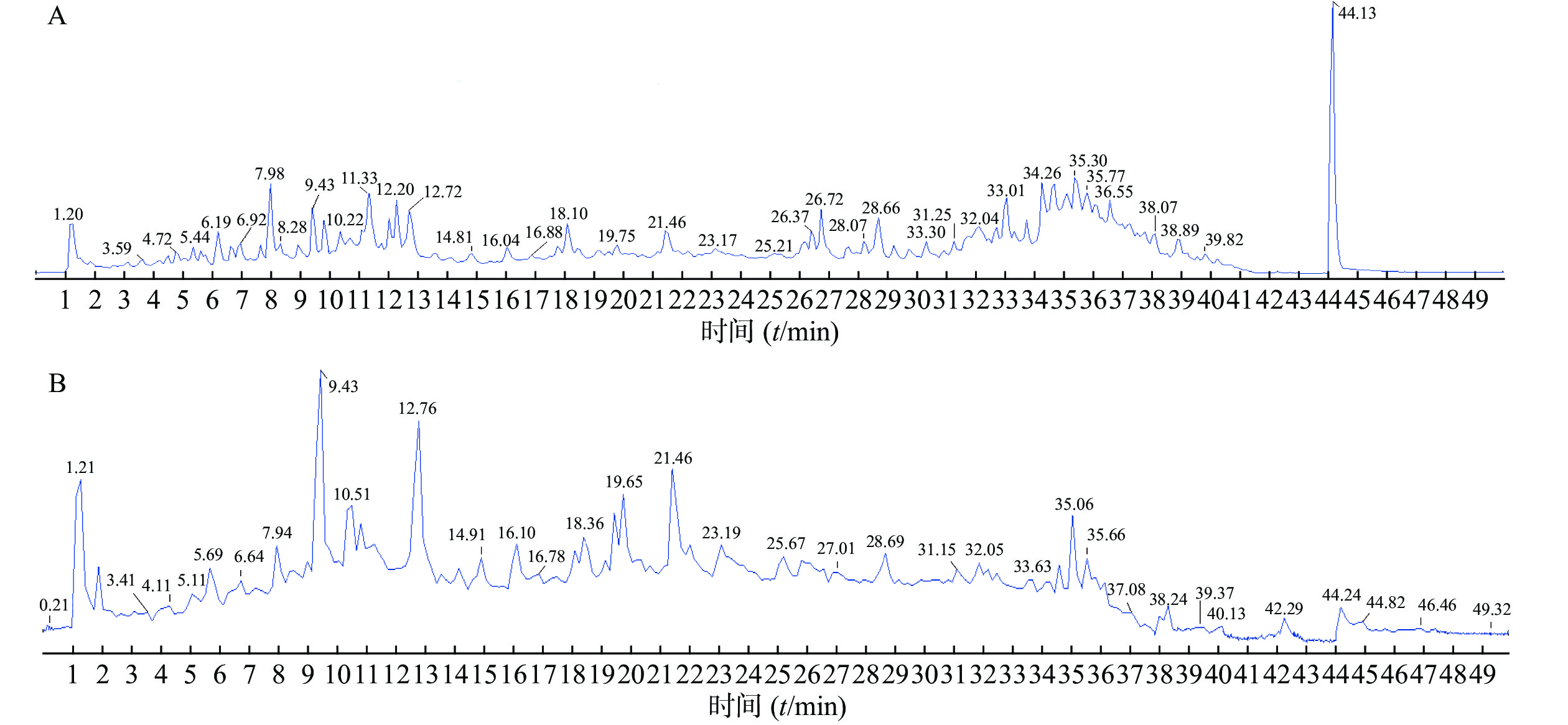
精密吸取对照品溶液和供试品溶液2 μl,采用“2.1”项下的色谱与质谱条件对样品进行分析,通过正、负离子全扫描,获得正、负离子模式下的总离子流图,见图1。
通过与对照品比对,分子离子峰质谱数据解析,与参考文献比对,共鉴定出三色片醇提物乙酸乙酯部位42个化合物,结果见表2。
表 2 三色片提取物中各成分主要碎片离子及谱峰归属化合物
编号tR/min 分子式 理论值(m/z) 模式 实测值(m/z) 误差(×10−6) 碎片离子(m/z) 化合物名称 参考文献 1 3.54 C7H6O3 139.039 0 [M+H]+ 139.039 4 3.0 121.028 7 原儿茶醛 [10] 2 4.76 C21H27N3O3 370.212 5 [M+H]+ 370.214 3 4.8 249.124 6,160.112 6,95.013 3,166.086 6,100.076 2,91.054 8 南蛇藤糠酰胺碱 [11] 3 6.63 C22H22O10 447.128 6 [M+H]+ 447.130 4 4.2 285.077 5,270.053 5,253.050 8,225.055 6,137.023 5 毛蕊异黄酮-7-O-β-D-葡萄糖苷 [12] 4 7.04 C23H29N3O2 380.233 3 [M+H]+ 380.235 1 4.8 176.106 9,160.112 6,105.033 8,100.076 5 苯代南蛇碱 [11] 5 9.41 C9H10O5 197.045 6 [M-H]− 197.044 7 −4.2 179.038 3,135.044 3 丹参素 [10] 6 9.42 C9H8O4 179.035 0 [M-H]− 179.034 2 4.7 135.044 8 咖啡酸 [13] 7 11.16 C16H12O4 431.133 7 [M+H]+ 431.136 3 6.1 269.082 6,253.050 3,225.055 5,213.091 7,197.060 2,136.014 6,118.041 7 芒柄花苷 [12] 8 12.71 C16H12O5 285.075 8 [M+H]+ 285.077 4 5.7 270.053 4,253.050 3,225.055 3,137.023 5 毛蕊异黄酮* [12] 9 12.76 C17H16O5 301.107 1 [M+H]+ 301.109 0 6.4 167.070 8,152.047 3,147.043 2,105.034 0,123.043 3 astrapterocarpan [12] 10 13.06 C20H24O6 361.164 6 [M+H]+ 361.166 5 5.5 269.154 3,227.108 3,185.096 9,157.101 7,129.070 3,91.054 9 雷公藤甲素* [14-15] 11 13.92 C17H14O6 315.086 3 [M+H]+ 315.088 1 5.6 300.064 7,243.065 5,167.034 2 熊竹素 [16] 12 18.21 C18H12O7 341.065 6 [M+H]+ 341.066 9 3.9 295.060 7,277.050 9,249.056 0 丹酚酸G [17] 13 19.76 C26H20O10 491.098 4 [M-H]− 491.097 0 −2.9 311.054 9,293.044 6,267.064 6,135.044 7 丹酚酸C [18] 14 21.46 C16H12O4 269.080 8 [M+H]+ 269.082 7 4.1 253.015 3,237.052 6,225.055 5,213.092 3,136.015 9,118.041 7,197.060 2 芒柄花素 [12] 15 22.57 C36H45NO17 764.276 0 [M+H]+ 764.278 3 2.9 746.276 1,686.246 3,644.235 1,206.081 7,188.070 9,178.086 5 aquifoliunine E-Ⅲ [14] 16 23.45 C20H22O6 359.148 9 [M+H]+ 359.150 7 4.9 267.138 0,225.019 5,183.079 9,128.061 8,91.054 3 雷公藤内酯酮* [19] 17 24.01 C38H47NO19 822.281 5 [M+H]+ 822.284 1 3.2 804.275 8,204.066 2,176.071 4 alatusinnine [20] 18 25.01 C39H45NO19 832.265 9 [M+H]+ 832.269 0 3.8 804.273 3,194.081 9,176.071 2 hypoglaunine E [11] 19 26.14 C41H68O14 829.458 0 [M+COOH]− 829.460 7 3.3 783.457 9,621.404 3,489.357 2 黄芪甲苷* [14] 20 28.19 C38H47NO18 806.286 6 [M+H]+ 806.290 3 3.8 788.279 5,686.247 0,206.082 1, 178.086 5 雷公藤定宁 E [20] 21 28.65 C39H45NO18 816.271 0 [M+H]+ 816.273 9 3.6 798.261 9,756.250 9,206.081 3,178.086 1,160.075 2 1-去乙酰基雷公藤吉碱 [11] 22 28.70 C43H70O15 871.468 6 [M+COOH]− 871.470 8 2.5 825.470 2,765.448 2,489.356 8 黄芪皂苷Ⅱ [13] 23 29.09 C41H47NO20 874.276 4 [M+H]+ 874.278 5 2.3 856.269 2,846.282 9,828.272 3,674.245 1,204.065 6,176.070 7 雷公藤春碱 [11] 24 29.72 C38H47NO18 806.286 6 [M+H]+ 806.291 2 3.8 788.280 4,686.247 4,206.082 4 peritassine A [20] 25 30.24 C43H70O15 871.468 6 [M+COOH]− 871.470 3 2.5 825.464 1,765.440 5 异黄芪皂苷Ⅱ
(异构体1)[16] 26 30.89 C19H16O4 309.112 1 [M+H]+ 309.114 2 6.7 281.667 0,263.106 0,235.076 7 丹参醛 [21] 27 31.58 C43H70O15 871.468 6 [M+COOH]− 871.470 8 2.5 825.470 2,765.448 0 异黄芪皂苷Ⅱ
(异构体2)[13] 28 32.12 C21H20O4 337.143 4 [M+H]+ 337.142 5 −2.7 309.686 6 丹参新醌丁 [10] 29 32.16 C43H49NO19 884.297 2 [M+H]+ 884.299 7 2.8 856.304 5,674.246 0,204.663 0,176.071 2 雷公藤定碱 [14] 30 32.96 C45H72O16 913.479 1 [M+COOH]− 913.482 4 3.5 867.481 7,825.469 8,807.464 3,765.450 6 黄芪皂苷Ⅰ [16] 31 32.99 C41H47NO19 858.281 5 [M+H]+ 858.285 4 4.6 840.275 7,798.263 8,746.269 1,738.243 5,686.248 0,206.082 5,178.087 1 雷公藤晋碱 [20] 32 33.04 C38H47NO18 806.286 6 [M+H]+ 806.289 7 3.8 788.278 3,686.244 4,206.082 1,728.257 0 卫矛碱 [20] 33 33.70 C45H72O16 913.479 1 [M+COOH]− 913.484 7 3.8 867.478 5,825.283 5,807.458 4,765.432 6 异黄芪皂苷Ⅰ
(异构体1)[16] 34 34.60 C45H72O16 913.479 1 [M+COOH]− 913.483 3 3.8 867.477 8,825.282 1,807.456 4,765.443 2 异黄芪皂苷Ⅰ
(异构体2)[16] 35 34.92 C46H49NO22 968.281 9 [M+H]+ 968.286 3 4.5 856.677 0,838.257 4,684.228 8,204.065 6,178.070 8 雷公藤素B [20] 36 35.01 C43H49NO18 868.302 2 [M+H]+ 868.304 6 2.7 868.364 0,850.295 8,746.268 9,686.247 6,206.082 4,178.087 1 雷公藤次碱 [20] 37 35.02 C41H47NO17 826.291 7 [M+H]+ 826.295 1 4.2 808.285 3,748.264 0,206.082 2,178.086 8 tripterygiumine Ⅰ [20] 38 35.49 C19H20O3 297.148 5 [M+H]+ 297.145 0 4.8 251.144 0,279.139 3,254.054 9,268.110 5,282.126 3 隐丹参酮 [10,17] 39 35.70 C20H28O2 299.201 7 [M-H]− 299.199 6 −6.7 283.168 2,213.090 8,201.916 0, 雷酚萜 [22] 40 35.86 C48H51NO18 930.317 9 [M+H]+ 930.321 3 3.7 912.308 7,310.111 0,206.081 8,188.071 2,178.086 5,105.033 6 ebenifoline E-Ⅱ [20] 41 36.81 C19H18O3 295.132 9 [M+H]+ 295.134 9 4.0 277.124 3,249.127 5,266.095 3,262.097 7,280.109 9 丹参酮Ⅱ A* [10,18,23] 42 37.49 C19H22O2 283.169 3 [M+H]+ 283.169 3 0 265.098 1,240.032 2,223.106 7,195.095 8,181.101 1 丹参新酮 [17,21,24] 注:*表示与对照品鉴定的化合物。 3.2.1 黄酮类化合物结构解析
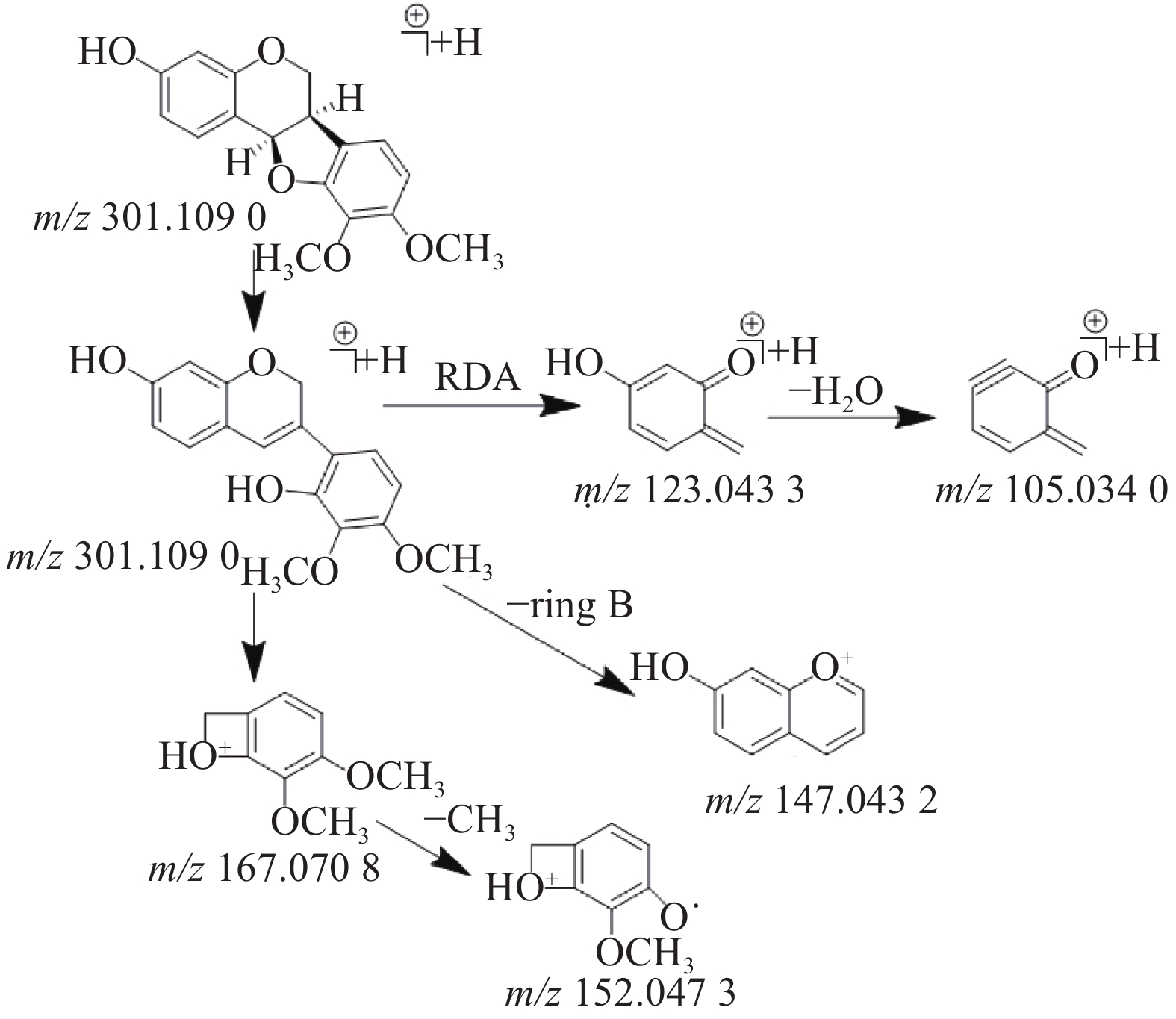
在乙酸乙酯部位中共鉴定出6个黄酮类化合物,其中4个黄酮苷元和2个黄酮苷,苷元为黄酮、异黄酮和紫檀烷,该类化合物在正离子模式下具有较好的响应。二级质谱中黄酮苷元易发生中性丢失,形成[M+H-H2O]+、[M+H-CO]+、[M+H-CH3]+等碎片离子,如在化合物8的二级质谱中可见m/z 270.053 4和m/z 253.050 3,则为m/z 285.077 4分别脱去-CH3和CH3OH形成的[M+H-CH3]+和[M+H-CH3OH]+碎片离子峰,m/z 225.055 3是m/z 253.050 3脱去1分子的CO形成的碎片离子峰,通过对照品的保留时间和参考文献[12]质谱数据比对确定化合物8为毛蕊异黄酮,m/z 137.023 5的碎片离子峰为异黄酮母核C环发生RDA裂解所产生。黄酮苷类易脱去糖基形成较强的分子离子峰,如化合物3(m/z 447.130 4)的二级质谱脱去糖基形成m/z 285.077 5的分子离子峰,并与化合物8(m/z 285.077 4)的二级质谱图非常相似,说明化合物3和化合物8在结构上是相似的,但化合物3的分子量多了162(C6H10O5),通过数据库比对和参考文献[12]推测化合物3则为毛蕊异黄酮-7-O-β-D-葡萄糖苷。化合物9(m/z 301.109 0)通过数据库比对发现两种候选化合物分别为astrapterocarpan和astraisoflavan,二级质谱中主要碎片离子峰为C环裂解产生的含A环和B环片段的碎片离子,其中,m/z 167.070 8为含B环的碎片离子峰且为基峰,进一步脱甲基形成m/z 152.047 3,m/z 123.043 3为含A环的碎片离子峰,进一步脱水形成m/z 105.034 0,m/z 147.043 2为母离子m/z 301.109 0脱去B环形成的碎片离子峰,根据m/z 167.070 8的碎片离子峰为基峰和含有m/z 147.043 2的碎片离子峰这两个特征,结合参考文献[12]的质谱数据,推测该化合物为astrapterocarpan,其相关裂解途径见图2。
3.2.2 三萜皂苷类化合物结构解析
在乙酸乙酯部位中鉴定出7个三萜皂苷类化合物,在负离子模式下均具有较好的响应,一级质谱中产生[M+COOH]−的准分子离子峰,二级质谱中产生较强的[M-H]-碎片离子峰和脱去糖基的较弱的分子离子峰。化合物19在负离子模式下产生的准分子离子峰为[M+COOH]−(m/z 829.458 0),二级质谱中产生m/z 783.457 9[M-H]−峰,脱去1分子六碳糖(C6H10O6)形成m/z 621.404 3的碎片离子峰,m/z 489.357 2则为m/z 621.404 3进一步脱去1分子五碳糖(C5H6O5)后形成的苷元碎片离子峰,推测其苷元为9,19-环阿尔廷烷,通过对照品的保留时间,参考文献[13]的离子碎片比对确定该化合物为黄芪甲苷。
3.2.3 生物碱类化合物结构解析
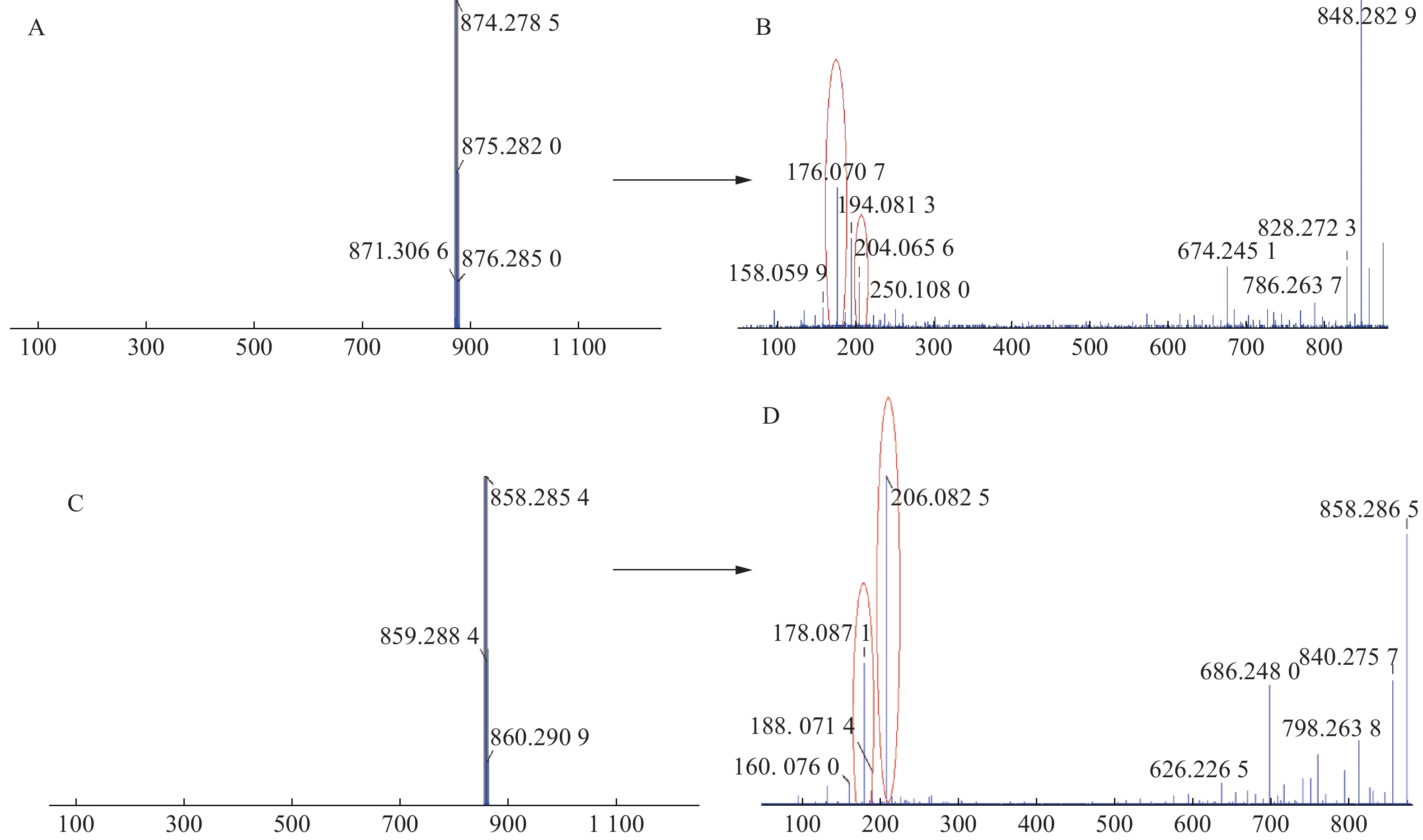
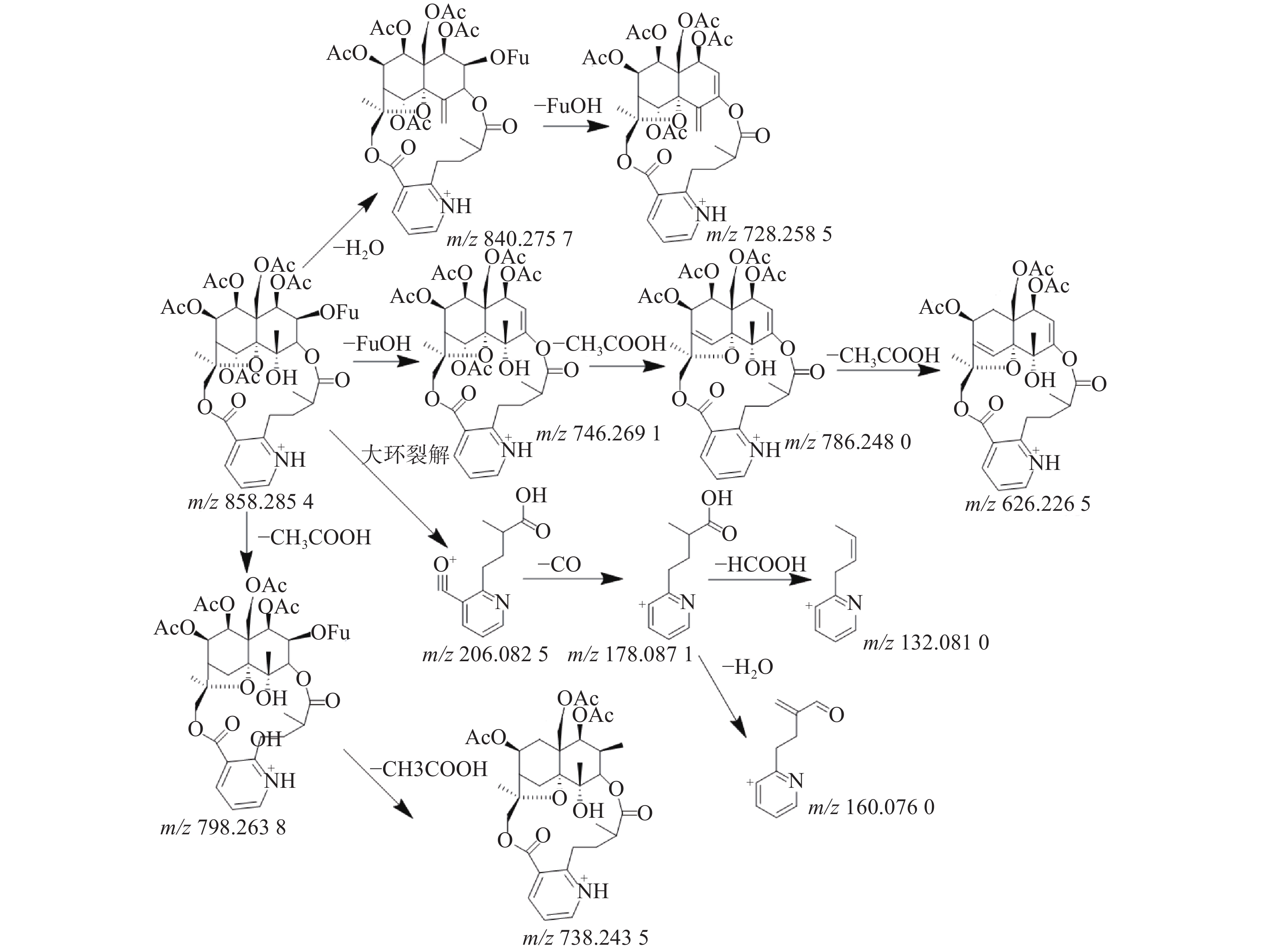
三色片提取物中共鉴定出16个生物碱类化合物,均来自雷公藤药材,在正离子模式下具有较好的响应,一级质谱中产生[M+H]+的准分子离子峰,二级质谱发现该类型的化合物容易脱去H2O、CO和CH3COOH等中性小分子而产生碎片离子峰,多数生物碱含有吡啶二羧酸部位的碎片离子峰。如化合物23在正离子模式下产生m/z 874.278 5的准分子离子峰,二级质谱中产生脱去1分子CO的m/z 846.282 9的基峰,脱去1分子H2O的m/z 856.2692的碎片离子峰和脱去1分子HCOOH的m/z 828.272 3的碎片离子峰,m/z 674.245 1峰为m/z 846.2829脱去C5H4O3侧链和CH3COOH形成的碎片离子峰,m/z 204.065 6峰为大环开裂产生的吡啶二羧酸部分脱水产生的碎片离子,该离子进一步脱羧形成m/z 176.070 7的碎片离子,通过数据库和参考文献[11]质谱数据的比对,推测化合物23为雷公藤春碱。化合物31在正离子模式下产生m/z 858.285 4的准分子离子峰,二级质谱中产生脱去1分子H2O的m/z 840.275 7的碎片离子峰,准分子离子峰脱去1分子CH3COOH形成较强的m/z 798.263 8峰,在进一步脱去1分子CH3COOH形成738.243 5峰,准分子离子峰m/z 858.285 4脱去FuOH(C5H4O3)侧链形成的m/z 746.269 1的碎片离子峰,再进一步脱去1分子CH3COOH,形成m/z 686.248 0的碎片离子,m/z 206.082 5峰为大环开裂产生的吡啶二羧酸部分脱水产生的碎片离子,该离子进一步脱羧形成m/z 178.087 1的碎片离子,通过数据库和参考文献[20]质谱数据的比对,推测化合物31为雷公藤晋碱。雷公藤晋碱中吡啶二羧酸部分较雷公藤春碱中少一个羟基,故其易产生m/z 206.0825的碎片离子峰,并通过脱羧产生m/z 178.087 1峰。两种化合物的质谱图见图3。以雷公藤晋碱为例,解析此类化合物的裂解规律,见图4。因此得出吡啶二羧酸部分含有羟基的生物碱会产生m/z 204系列的特征碎片离子峰,不含羟基的生物碱则产生m/z 206系列的特征碎片离子峰。
3.2.4 萜类化合物结构解析
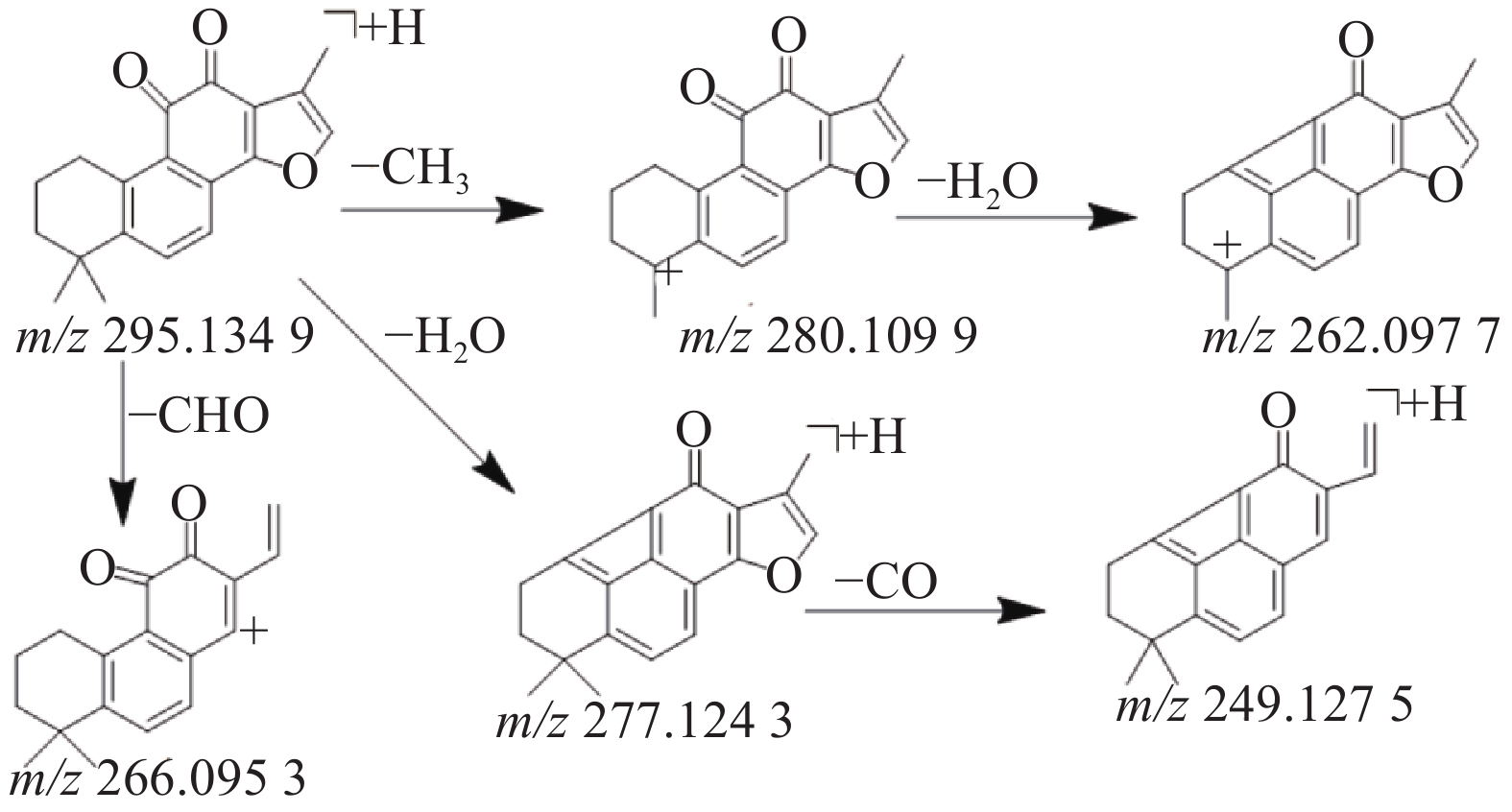
本研究共鉴定出8种萜类化合物,其中源于丹参药材中的5种萜类成分,丹参中的萜类化合物因其结构中主要含有羟基,羰基等取代基,所以质谱碰撞中主要丢失H2O,CO和-CH3等中性分子,产生一系列的碎片离子峰。化合物41在正离子模式下产生m/z 295.134 9的[M+H]+准分子离子峰,二级质谱中产生脱去1分子甲基形成的m/z 280.1099的碎片离子峰,在此基础上有丢失1分子水形成m/z 262.097 7峰,准分子离子峰脱去1分子H2O或脱去1个-CHO形成m/z 277.124 3峰或m/z 266.095 3峰,m/z 249.127 5峰是m/z 277.124 3脱去1分子H2O形成的碎片峰,通过对照品的保留时间和参考文献[10,18,23]数据比对,鉴定该化合物为丹参酮Ⅱ A,其质谱裂解规律见图5。
来源于雷公藤药材中的3种二萜类成分,该类化合物的二级质谱中出现一系列的脱水、脱CO和异丙基等碎片离子峰。化合物11在正离子模式下产生m/z 361.166 5的准分子离子峰,脱去2分子H2O和2分子CO形成m/z 269.154 3的碎片离子峰,m/z 227.108 3为m/z 269.154 3脱去1分子CH2CHCH3形成的碎片离子,其进一步脱1分子H2O和HCHO形成m/z 185.096 9的碎片离子,通过对照品比对和参考文献[14-15]的质谱数据,确定化合物10为雷公藤甲素。化合物18在正离子模式下产生m/z 359.148 9的准分子离子峰,脱去2分子H2O和2分子CO形成m/z 267.138 0的碎片离子峰,m/z 225.019 5为m/z 267.138 0脱去1分子CH2CHCH3形成的碎片离子,其进一步脱1分子H2O和HCHO形成m/z 183.0799的碎片离子,通过对照品比对和参考文献[19]的质谱数据,确定化合物16为雷公藤内酯酮。化合物39在负离子模式下产生m/z 299.199 6 的准分子离子峰,二级质谱中产生m/z 283.168 2的碎片离子, 提示为丢失1个-CH3后形成双键产生的碎片离子峰,A环发生RDA裂解产生m/z 213.090 8的碎片离子峰,通过数据库比对和参考文献[22]的质谱数据,推测化合物39为雷酚萜。
3.2.5 酚酸类化合物结构解析
在正负离子模式下共鉴定出乙酸乙酯部位中5种酚酸类成分,均来自于丹参药材,参考文献[18]报道的丹参中酚酸类成分的裂解规律发现,酚酸类化合物主要含有羰基、羧基和羟基,所以在质谱碰撞中易丢失CO、H2O和CO2的中性碎片;丹参素和咖啡酸作为基本母核而其他的水溶性酚酸类化合物大多数为这两者的聚合或缩合产物,主要为缩酚酸类的成分,在质谱碰撞中易丢失[M-H-180]−和[M-H-198]−中性碎片;含有羧基的单体化合物在负离子模式下会产生135[C8H7O2]−和179[C9H7O4]−的特征性碎片。化合物5中,在负离子模式下产生m/z 197.044 7的[M-H]−准分子离子峰,二级质谱进一步产生丢失1分子H2O和1分子CO2,形成的m/z 179.038 3和m/z 135.044 3的碎片离子峰,推测出结构中含有羧基,结合其精确分子量和参考文献[10]质谱数据,推测该化合物为丹参素。化合物12中,负离子模式下产生m/z 491.097 0的[M-H]−准分子离子峰,二级质谱中产生m/z 311.054 9和m/z 293.044 6的碎片离子峰,分别为[M-H-180]−和[M-H-198]−, m/z 267.064 6峰为m/z 311.054 9脱去1分子CO2所产生,根据m/z 135.044 7峰推测结构中含有羧基,结合其精确分子量和参考文献[18]质谱数据的比较,推测该化合物为丹酚酸C。化合物1中,正离子模式下给出m/z 139.039 4的[M+H]+准分子离子峰,脱去1分子H2O形成m/z 121.028 7的碎片离子峰,通过数据库比对和参考文献[10],推测该化合物1为原儿茶醛。化合物13中,在正离子模式下产生m/z 341.066 9的[M+H]+准分子离子峰,脱去1分子CO2形成m/z 295.060 7的碎片离子峰,m/z 277.050 9和m/z 249.056 0的碎片离子峰是m/z 295.060 7峰分别脱去1分子H2O和1分子CO2形成的,通过数据库比对和参考文献[17]质谱数据,推测该化合物为丹酚酸G。
4. 讨论
4.1 色谱与质谱条件考察
本实验流动相考察了乙腈-水系统和甲醇-水系统,结果乙腈-水系统中化合物的分离度较好,加入甲酸可以改善峰形,有助于化合物的离子化,提高质谱的响应,最终选择乙腈-0.1%甲酸水系统作为本次研究的流动相。
4.2 化学成分的定性分析
据以往文献中三色片各化学成分的研究报道,收集各药材的主要化学成分的精确分子量,碎片离子峰等信息,建立相应的化学成分数据库。通过数据库比对,对照品保留时间及参考文献中质谱数据鉴定三色片醇提物乙酸乙酯部位的化学成分。本研究共鉴定出42个化合物,其中5个是通过对照品鉴定得出,对无对照品的化合物,通过质谱的裂解特征及参考文献进行结构表征,对同分异构体应结合其在液相色谱中化合物的保留时间及质谱行为,综合对其定性鉴别。
三色片醇提物的乙酸乙酯部位具有较强的抗补体活性,本研究采用UPLC-Q-TOF-MS法对其中的化学成分进行结构表征,结果发现该部位主要含有生物碱类,萜类,黄酮和酚酸类等化学成分。其中以来源于雷公藤药材中极性中等的生物碱类成分含量较多,这与三色片提取物的制备工艺有关,三色片中雷公藤药材采用乙醇加热回流提取的方式,而黄芪和丹参药材采用水提取醇沉淀的方式。此外,先前的研究发现广藿香中的黄酮和萜类化合物对旁路途径的补体激活具有明显的抑制作用,紫花地丁中的生物碱类成分对旁路途径也有抑制作用(AP50=0.22~0.50 g/L), 牡丹皮和毛七公的抗补体活性成分研究中发现酚羟基决定抗补体活性的存在与否,没食子酰基可改善抗补体活性,甲基则对抗补体活性不利[25-27]。通过本次研究对三色片醇提物的乙酸乙酯部位的化学成分进行了初步表征,为阐明三色片的药效物质基础提供参考依据。研究的不足之处在于,仍有部分化学成分尚未定性鉴定,含量较高的单体成分未进行体外抗补体活性的测定,未来将通过中药化学的方法获得含量较高的单体成分,并进行结构鉴定和抗补体活性测定。
-
图 2 水母胶原蛋白纯化过程SDS-PAGE电泳图
M.蛋白Marker;1~2.分子量分别是5万、10万透析袋处理;3~5.分别是0.9 mol/L→0.9 mol/L→0.9 mol/L、0.9 mol/L→0.9 mol/L→2.0 mol/L、2.0 mol/L→0.9 mol/L→0.9 mol/L NaCl的盐析处理;6~7.分子量分别是10万和5万的超滤处理;8.水母胶原蛋白粗提物
表 1 水母胶原蛋白提取正交实验因素水平表
水平 因素 A加酶量(%) B乙酸浓度(mol/L) C料液比 D反应时间(h) 1 0.1 0.3 1∶1 24 2 0.2 0.4 1∶2 48 3 0.3 0.5 1∶3 72  下载: 导出CSV
下载: 导出CSV
表 2 水母胶原蛋白提取正交实验结果
水平 因素 A 加酶量(%) B 乙酸浓度(mol/L) C料液比(g/ml) D时间(h) 胶原蛋白得率(%) 实验1 0.1 0.3 1∶1 24 17.8±0.66 实验2 0.1 0.4 1∶2 48 23.2±0.94 实验3 0.1 0.5 1∶3 72 21.7±0.85 实验4 0.2 0.3 1∶2 72 26.7±0.58 实验5 0.2 0.4 1∶3 24 19.0±0.86 实验6 0.2 0.5 1∶1 48 24.2±0.76 实验7 0.3 0.3 1∶3 48 26.9±0.61 实验8 0.3 0.4 1∶1 72 31.1±0.50 实验9 0.3 0.5 1∶2 24 24.3±0.75 K1 20.9 23.8 24.3 20.4 K2 23.3 24.4 24.7 24.8 K3 27.4 23.4 22.5 26.5 R 6.53 1.03 2.20 6.13 因素主次 A>D>C>B 最佳组合 A3B2C2D3
下载: 导出CSV
表 3 胶原蛋白X射线衍射峰对应的d值
组别 衍射峰 2θ/(°) d值(nm) 水母胶原蛋白 峰1 7.1 1.244 峰2 21.3 0.418 峰3 30.2 0.295 牛胶原蛋白 峰1 7.2 1.226 峰2 20.3 0.437 峰3 30.1 0.296
下载: 导出CSV
表 4 二种胶原蛋白中不同氨基酸组分的百分比
氨基酸名称 氨基酸占比(%) 水母胶原蛋白 牛胶原蛋白 天冬氨酸Asp 7.83 4.57 苏氨酸Thr 3.55 1.73 丝氨酸Ser 4.89 3.19 谷氨酸Glu 9.98 7.27 甘氨酸Gly 34.82 34.37 丙氨酸Ala 8.36 12.46 缬氨酸Val 2.74 2.27 蛋氨酸Met 0.21 0.01 异亮氨酸Ile 1.88 1.22 亮氨酸Leu 3.09 2.64 酪氨酸Tyr 0.11 0.27 苯丙氨酸Phe 0.9 1.38 赖氨酸Lys 2.96 2.15 组氨酸His 0.29 0.48 精氨酸Arg 3.87 5.3 脯氨酸Pro 8.16 12.43 羟脯氨酸Hyp 6.33 8.26 合计 100 100 注:氨基酸分析采用氨基酸残基数占100个氨基酸残基的比例
下载: 导出CSV
-
[1] WIDDOWSON J P, PICTON A J, VINCE V, et al. In vivo comparison of jellyfish and bovine collagen sponges as prototype medical devices[J]. J Biomed Mater Res B Appl Biomater,2018,106(4):1524-1533. doi: 10.1002/jbm.b.33959 [2] SILVA T H, MOREIRA-SILVA J, MARQUES A L, et al. Ma- rine origin collagens and its potential applications[J]. Mar Drugs,2014,12(12):5881-5901. doi: 10.3390/md12125881 [3] LI P H, LU W C, CHAN Y J, et al. Extraction and characterization of collagen from sea cucumber (Holothuria cinerascens) and its potential application in moisturizing cosmetics[J]. Aquaculture,2020,515:734590. doi: 10.1016/j.aquaculture.2019.734590 [4] KHONG N M, YUSOFF F M, JAMILAH B, et al. Nutritional composition and total collagen content of three commercially important edible jellyfish[J]. Food Chem,2016,196:953-960. doi: 10.1016/j.foodchem.2015.09.094 [5] COENTRO J Q, CAPELLA-MONSONÍS H, GRACEFFA V, et al. Collagen quantification in tissue specimens[J]. Methods Mol Biol,2017,1627:341-350. [6] 李玉芬, 郑明星, 朱凡, 等. 海蜇胶原蛋白的制备及理化性质研究[J]. 福州大学学报(自然科学版), 2018, 46(2):286-294. [7] ZHANG J J, DUAN R, HUANG L, et al. Characterization of acid-soluble and pepsin-solubilised collagen from jellyfish (Cyanea nozakii Kishinouye)[J]. Food Chem,2014,150:22-26. doi: 10.1016/j.foodchem.2013.10.116 [8] SUN L L, LI B F, SONG W K, et al. Characterization of Pacific cod (Gadus macrocephalus) skin collagen and fabrication of collagen sponge as a good biocompatible biomedical material[J]. Process Biochem,2017,63:229-235. doi: 10.1016/j.procbio.2017.08.003 [9] 姚行行, 郭妍, 庄永亮. 云南鲷鱼骨胶原蛋白的制备及其理化性质[J]. 食品科学, 2018, 39(13):35-40. doi: 10.7506/spkx1002-6630-201813006 [10] DE CHEN J, LI L, YI R Z, et al. Extraction and characterization of acid-soluble collagen from scales and skin of tilapia (Oreochromis niloticus)[J]. LWT - Food Sci Technol,2016,66:453-459. doi: 10.1016/j.lwt.2015.10.070 [11] AB AZIZ N A, SALIM N, ZAREI M, et al. Extraction, anti-tyrosinase, and antioxidant activities of the collagen hydrolysate derived from Rhopilema hispidum[J]. Prep Biochem Biotech- nol,,2020:1-10. [12] RASTIAN Z, PÜTZ S, WANG Y, et al. Type I collagen from jellyfish Catostylus mosaicus for biomaterial applications[J]. ACS Biomater Sci Eng,2018,4(6):2115-2125. doi: 10.1021/acsbiomaterials.7b00979 [13] NOORZAI S, VERBEEK C J R, LAY M C, et al. Collagen extraction from various waste bovine hide sources[J]. Waste Biomass Valor,2020,11(11):5687-5698. doi: 10.1007/s12649-019-00843-2 [14] LIN X H, CHEN Y Y, JIN H X, et al. Collagen extracted from bigeye tuna (Thunnus obesus) skin by isoelectric precipitation: physicochemical properties, proliferation, and migration activi- ties[J]. Mar Drugs,2019,17(5):E261. doi: 10.3390/md17050261 [15] SONG W K, LIU D, SUN L L, et al. Physicochemical and biocompatibility properties of type I collagen from the skin of Nile tilapia (Oreochromis niloticus) for biomedical applications[J]. Mar Drugs,2019,17(3):E137. doi: 10.3390/md17030137 [16] ZHANG X, CHEN Y R, ZHAO Y L, et al. Type I collagen or gelatin stimulates mouse peritoneal macrophages to aggregate and produce pro-inflammatory molecules through upregulated ROS levels[J]. Int Immunopharmacol,2019,76:105845. doi: 10.1016/j.intimp.2019.105845 [17] RODRÍGUEZ F, MORÁN L, GONZÁLEZ G, et al. Collagen extraction from mussel byssus: a new marine collagen source with physicochemical properties of industrial interest[J]. J Food Sci Technol,2017,54(5):1228-1238. doi: 10.1007/s13197-017-2566-z [18] WALTER M N, DEHSORKHI A, HAMLEY I W, et al. Supra-molecular assembly of a lumican-derived peptide amphiphile enhances its collagen-stimulating activity[J]. Biomater Sci,2016,4(2):346-354. doi: 10.1039/C5BM00428D [19] CHINH N T, MANH V Q, TRUNG V Q, et al. Characterization of collagen derived from tropical freshwater carp fish scale wastes and its amino acid sequence[J]. Nat Prod Commun,2019,14(7):1-12. [20] BOU-GHARIOS G, ABRAHAM D, DE CROMBRUGGHE B. Type I collagen structure, synthesis, and regulation[M]//Principles of Bone Biology. Amsterdam: Elsevier, 2020: 295-337. [21] ASADUZZAMAN A K M, GETACHEW A T, CHO Y J, et al. Characterization of pepsin-solubilised collagen recovered from mackerel (Scomber japonicus) bone and skin using subcritical water hydrolysis[J]. Int J Biol Macromol,2020,148:1290-1297. doi: 10.1016/j.ijbiomac.2019.10.104 [22] ELANGO J, LEE J W, WANG S J, et al. Evaluation of differentiated bone cells proliferation by blue shark skin collagen via biochemical for bone tissue engineering[J]. Mar Drugs,2018,16(10):E350. doi: 10.3390/md16100350 [23] COELHO R C G, MARQUES A L P, OLIVEIRA S M, et al. Extraction and characterization of collagen from Antarctic and Sub-Antarctic squid and its potential application in hybrid scaffolds for tissue engineering[J]. Mater Sci Eng C Mater Biol Appl,2017,78:787-795. doi: 10.1016/j.msec.2017.04.122 [24] KRISHNAN S, PERUMAL P. Preparation and biomedical cha- racterization of jellyfish (Chrysaora Quinquecirrha) collagen from Southeast Coast of India[J]. Int J Pharm Pharm Sci,2013,5(3):698-701. [25] WICHUDA J, SUNTHORN C, BUSARAKUM P. Comparison of the properties of collagen extracted from dried jellyfish and dried squid[J]. Afr J Biotechnol,2016,15(16):642-648. doi: 10.5897/AJB2016.15210 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5931
- HTML全文浏览量: 2342
- PDF下载量: 99
- 被引次数: 0

