

 下载:
下载:



-
念珠菌为条件致病真菌,可寄居于人体的不同部位,当人体局部或全身免疫力下降时,其致病性可造成个体浅表或全身感染。白念珠菌(Candidaalbicans)是念珠菌的一种,是引发侵袭性感染的最主要致病菌[1]。近年来,由于抗真菌药物的大量使用,白念珠菌的耐药性逐渐增加。常规抗菌药对白念珠菌的抑制效果明显降低。
相对于单药而言,多种药物联合使用时往往能通过药效互补,起到协同作用,增强抑菌效果[2-3]。联合用药提高了抑菌效果,患者的治疗周期缩短,治疗成本可进一步降低。因此,研究抗白念珠菌药物联合疗法对于治疗白念珠菌感染非常有意义。
一般来说,联合疗法会同时使用抗真菌药和增效剂。目前经典的抗真菌药物有唑类化合物(如氟康唑)、多烯类(如两性霉素B)和棘白菌素类(如卡泊芬净)等,这些药物可直接抑制或杀伤白念珠菌。氯法齐明、法尼醇本身对真菌没有直接抑制作用,但可作为增效剂辅助增强抗真菌药物治疗效果,增强耐药菌的敏感性。近年来,光动力学、声动力学疗法等新兴的技术应用于抗真菌的治疗,同时传统的抗真菌物质在现代技术的改进下也能发挥特有的优势,比如将银制成纳米银颗粒用于抑制白念珠菌增殖。本文将针对白念珠菌尤其是耐药菌的杀伤或抑制作用,聚焦于传统抗真菌药物与声动力学/光动力学疗法、纳米银颗粒这几类抗菌方式,阐述与之相关的联合疗法。
-
氟康唑、酮康唑、咪康唑等唑类化合物是治疗白念珠菌感染的经典药物,具有毒性相对较低、制备技术成熟等优点。但随着临床治疗疗程变长以及使用频率的增多,白念珠菌的耐药性也越来越严重,单独使用唑类化合物的药效大大降低[4]。研究表明白念珠菌对唑类化合物耐药性主要由以下几种机制引起:唑类药物靶点ERG11过度表达降低了白念珠菌的敏感性;ERG11突变导致与唑类化合物的亲和力降低;细胞膜外排泵表达增加,将唑类药物排出细胞外;ERG3突变避免了氟康唑处理后有毒副产物在细胞内的积累;生物被膜物理隔离了白念珠菌胞体和抗真菌药物的接触[5]。尽管单独使用唑类药物应对耐药菌的效果并不理想,但作为传统的抗真菌药物,其作用机制较为明确,在与其他类型的抗真菌药物或疗法的协同作用下,仍发挥较强的抗真菌效果。
-
真菌生物被膜是白念珠菌产生耐药性的重要原因之一。有实验表明,高浓度的酪醇、法尼醇(合金欢醇)等群感效应分子与氟康唑联合使用,可以起到抗真菌被膜的效果,提高白念珠菌对药物的敏感性[6]。白念珠菌对氟康唑等药物耐药的另一个重要机制是其细胞上的外排泵蛋白。对白念珠菌转运泵活性的抑制可增加真菌胞内药物的浓度,以达到抑制或杀灭真菌的目的。研究发现,法尼醇、盐酸氨溴索具有抑制ATP结合转运蛋白的功能,与氟康唑联用可达到抑制氟康唑耐药菌的作用。法尼醇单独使用时还可以诱导氟康唑抗性白念珠菌产生细胞内活性氧,促使其发生凋亡[7]。
-
天然化合物在联合用药中也可以发挥重要作用,将其与常规药物结合可以成为应对耐药菌的有效策略。部分生物碱类,如以小檗碱及其衍生物为代表的异喹啉生物碱以及依波加因等吲哚生物碱,尽管在单独使用没有抗真菌功效,但当其与氟康唑联合使用时有克服白念珠菌对氟康唑耐药的效果[8-9]。还有一些化合物,例如,没食子酸、土槿皮乙酸,单独使用时即表现出抗真菌作用,与氟康唑联合使用可明显降低氟康唑的最低抑菌浓度[10-11]。此外,壳聚糖也具有一定的抗真菌效果,可以通过抑制染色质重塑复合物SAGA表达来影响白念珠菌细胞壁和细胞膜的形态[12]。氟康唑联合壳聚糖,有协同抗真菌活性[13]。得益于壳聚糖的细胞亲和性、生物降解性、低毒性、制备技术成熟等优势,氟康唑与壳聚糖的联合应用具有很大潜力。同时,中药方剂黄连解毒汤与氟康唑联用,通过抑制麦角甾醇相关基因,破坏白念珠菌生物被膜的形成,为抗真菌治疗提供了研究思路[14]。除此之外,一些皮质类固醇,例如地塞米松、布地奈德等药物,也能增强白念珠菌对氟康唑的敏感性[15-16]。
-
钙调磷酸酶在真菌细胞形态、耐药性等多个生理方面发挥着重要作用[17]。氟康唑与FK506、环孢素A等钙调磷酸酶抑制剂联合使用,可以有效降低白念珠菌对氟康唑的耐受性。一方面,氟康唑抑制白念珠菌麦角甾醇的合成,使钙调磷酸酶抑制剂更容易进入白念珠菌细胞内;另一方面,钙调磷酸酶抑制剂增强了氟康唑的药效,使其从抑菌效果转为杀菌效果。两者联用能破坏白念珠菌的生物被膜,增强了抗真菌效果[18]。
-
HSP90是一类与白念珠菌毒力和耐药性密切相关的分子伴侣,可以促进外排泵Cdr1的高表达,增强白念珠菌排出唑类药物,还可以通过一系列级联反应诱发白念珠菌对唑类耐受[19]。因此,抑制HSP90活性是提高白念珠菌对唑类化合物敏感性的有效途径之一。HSP90抑制剂Ganetespib可以下调氟康唑作用的靶酶ERG11和白念珠菌外排泵基因CDR1的表达,有抗生物被膜的效果,与氟康唑在体外联合使用时具有抗真菌活性[20]。除此之外,一些其它的HSP90抑制剂,如格尔德霉素,也可以和氟康唑起到协同作用[21]。针对真菌HSP90设计制备的人类基因重组抗体Mycograb与氟康唑的协同效果仅局限于敏感菌,但当其与两性霉素B联合使用时却能对部分耐药菌表现出一定的协同作用[22]。
-
两性霉素B是经典的多烯类抗真菌药物,具有杀菌活性,但对人体不良反应较多。两性霉素B可以与成熟真菌细胞的细胞壁和出芽产生的子细胞的质膜结合,破坏真菌细胞膜,从而发挥抗真菌效果[23]。相对唑类药物而言,两性霉素B耐药发生的情况较少,目前主要面临的问题在于两性霉素B的毒性较大,尤其是对肝肾功能不全的患者有较大的副作用。所以在选择与两性霉素B联合使用的药物时,既要考虑抑菌效果又要考虑药物毒性,在联合使用时最好是能够通过低剂量两性霉素B达到较好效果,以降低其毒性产生的影响。为了降低两性霉素B对人体的毒性,常用的是采用结合脂质体的方法,将两性霉素B包埋在脂质体试剂中。脂质体可以高效地介导两性霉素B与真菌结合,在减少两性霉素B副反应的同时,还提高了其转移至真菌细胞内的效率[24]。两性霉素B和5-氟胞嘧啶联合是临床常用方法,具有更好的抗真菌功效,而且相比于单独使用两性霉素B,两者联合可小幅度降低两性霉素B的毒性[30]。
一些具有抗真菌效果的化合物可能同时具有低毒性的优势。天然化合物丁香酚可以作为渗透酶抑制剂影响白念珠菌的生长形态并阻断Ca2+通道,将其与两性霉素B同时使用时能够引起白念珠菌细胞内活性氧含量的升高,线粒体损伤,胞质内细胞色素C含量升高,导致细胞损伤。相对于单独发挥抗菌效果的两性霉素B,该组合有效降低了两性霉素B的使用剂量,降低了毒副作用[25]。 1,3,4-噻二唑衍生物4-(5-methyl-1,3,4-thiadiazole-2-yl) benzene-1,3-diol(C1化合物)可与白念珠菌的细胞壁发生作用,影响其完整性,诱发形态紊乱。研究表明,C1化合物与两性霉素B显示出了强烈的协同作用,将100%抑制病原真菌所需的两性霉素B用量降低了10倍甚至几十倍,而且针对正常人皮肤成纤维细胞(NHDF)的细胞毒实验表明,高浓度的C1化合物并不会对其产生细胞毒效应[26]。
与其他非抗真菌药物联合使用以增强抗真菌效果的方法对两性霉素B同样适用。近年来,已经有团队研发了Simpotentin、Nectriatide、Polyketide glycosides phialotides等两性霉素B活性增强剂,临床前阶段的实验表明,这些两性霉素B活性增强剂可以有效地降低两性霉素B的用量[27-29]。
-
棘白菌素是一类β-1,3-D-葡聚糖合酶抑制剂,可以干扰白念珠菌细胞壁的形成,随着其临床上的广泛应用,有越来越多的耐药菌被分离出来。白念珠菌编码葡聚糖合酶亚基的FKS基因突变、真菌生物被膜的形成、应激适应等多种因素均可能引发棘白菌素耐药,通过联合疗法有可能解决耐药问题[31]。
-
尼可霉素Z是一种几丁质合酶抑制剂,同β-1,3-D-葡聚糖一样,几丁质也是真菌细胞壁的重要组成成分。当棘白菌素类药物抑制白念珠菌β-1,3-D-葡聚糖合成时,白念珠菌有可能通过上调几丁质的合成,减轻棘白菌素对细胞壁的损伤,降低对棘白菌素的敏感性[32]。如果在使用棘白菌素类药物卡泊芬净或米卡芬净的基础上同时使用尼可霉素Z,则可以同时抑制β-1,3-D-葡聚糖和几丁质的合成,有效抑制白念珠菌的生长[33]。
-
由于棘白菌素与唑类化合物抑制白念珠菌的机制不同,将两者联合使用增强对棘白菌素耐药菌的抑制作用。当棘白菌素与泊沙康唑联合使用时,对白念珠菌SC5314的分级抑菌浓度(FIC)<0.5,表明两者联合使用时具有体外协同抗真菌活性[34]。
-
氯法齐明是传统的抗分枝杆菌药物,尽管不是抗真菌药物,但研究发现,当氯法齐明与一些其他抗菌类药物联合使用时,具有广谱的抗真菌活性,因此有作为联合药物应用于抑制传统耐药菌增殖的潜力。有研究表明,氯法齐明分子的疏水结构可以嵌入到白念珠菌的膜双层的烃相中,通过非特异性膜扰动发挥对白念珠菌的抑制作用。当卡泊芬净与氯法齐明联合使用时,两者表现出了协同作用[35]。
-
一些小分子抗真菌肽和棘白菌素类药物联合使用,也有抑制白念珠菌生长的潜力。通常而言,这类多肽发挥抗菌作用的机制主要是抑制真菌生物膜的形成。植物防御素家族是抗真菌肽的丰富来源,对真菌病原体具有广谱的抗菌活性且通常对人类毒副作用较小。植物防御素HsAFP1在与卡泊芬净的协同作用下表现出了显著的抗白念珠菌生物被膜活性[36]。在应用于联合疗法时,可以考虑将抗真菌多肽通过一些处理来增强活性。酪氨酸复合物可以通过破坏白念珠菌生物膜、诱导内源性活性氧等效应发挥对白念珠菌的抑制作用,与卡泊芬净有明显的协同作用[37]。但由于其容易在水中形成高级结构,过度聚集,其活性会因此而受到影响。针对酪氨酸复合物易于纤维素稳定结合的特性,利用纤维素类制剂可以有效控制酪氨酸复合物的稳定性,增强其活性[38]。
-
声动力学和光动力学抗真菌疗法是一个新兴的技术,近年来,针对白念珠菌光动力学和声动力学疗法相关的研究也逐渐增多,在临床和实验都有其相关应用的报道。由于白念珠菌的耐药性与其对光动力学疗法抗性无直接的相关性,光动力学疗法有望成为应对白念珠菌耐药的有效手段之一[39]。在纳米颗粒、姜黄素类药物、甲苯胺蓝等光敏剂的作用下可以使用无害的可见光激活活性氧,从而达到破坏真菌生物被膜、杀伤真菌细胞的效果[40]。β葡聚糖酶、DNA酶等水解酶可以通过破坏白念珠菌生物被膜,使光敏剂更容易渗透到更深的生物膜层,加强光动力疗法的效果[41]。为了进一步提高光动力学疗法的抗真菌效果,可以使用其他抗真菌药物联合使用,有研究表明,相对于单独使用抗真菌的唑类药物,如咪康唑、氟康唑、伊曲康唑等,唑类药物和光动力学疗法联合使用时可以下调绝大部分白念珠菌菌株的活力[42-43]。
声动力学疗法主要基于低频和低强度超声,在与其他药物如两性霉素B的协同作用下,声动力学疗法比单独使用抗菌药物更有效,对白念珠菌活性、生物被膜活性以及其酶的活性均产生一定的影响[44-45]。
-
银和银盐用于灭菌已有很长的历史,近年来,随着纳米材料在生物学中的应用,纳米银的抗菌作用逐渐引起重视。纳米银颗粒主要是通过诱导细胞内活性氧的生成对白念珠菌产生杀伤作用,还可以诱导真菌耐药性和致病性相关的多个靶点改变,影响真菌细胞形态、膜流动性、菌丝形成等[46]。有实验表明,纳米银对多种真菌的混合生物被膜有抑制效果[47]。单独使用纳米银有一定的抗菌效果,不同的载体和不同的纳米银制备过程对其抗菌效果影响不同,相对于使用游离的纳米银颗粒,使用可生物降解的材料聚乳酸作为载体时的抗菌活性明显提高[48]。有研究表明,纳米银可以干扰白念珠菌对葡萄糖的摄取,当其与糖酵解抑制剂3-溴丙酮酸共同使用时,则可以进一步阻碍白念珠菌获取增殖所需的能量,针对能量层面表现出协同作用,促进白念珠菌的死亡[49]。
-
随着传统抗真菌药物的长期大量使用,白念珠菌的耐药性也逐渐增强。在探索新的抗真菌药物的同时,联合疗法也可以作为临床治疗中应对耐药白念珠菌的有效思路。抗菌药物和增强剂同时使用、新型抗菌技术配合药物等联合治疗手段,已经有了大量实验数据的支持,展现出了巨大的临床应用潜力。
在联合疗法的介导下,发挥抗菌作用的药物仍是抗菌主力,其他非抗菌药物和技术则有效地辅助了抗菌药物的疗效,通过破坏真菌生物被膜等方式发挥协同效应。在经典抗菌药物杀伤真菌的基础上,尝试联合以不同途径抑制、杀伤真菌或针对真菌耐药机制发挥作用的药物及手段,是寻找新的联合疗法的主要思路。
Research progress on drug combination therapy against Candida albicans
-
摘要: 白念珠菌是念珠菌属最常见的种类之一,是临床上重要的侵袭性念珠菌感染来源。由于传统抗真菌药物的广泛使用,临床已经分离出很多耐药白念珠菌,导致部分传统药物在临床治疗效果明显下降。同时,一些化合物自身的毒性限制了其临床应用。在这种背景下,联合疗法由于发挥了各种药物或手段的协同作用,有可能改进单独用药的不足,具有抑制白念珠菌生长的巨大潜力。Abstract: Candida albicans is one of the most common species of Candida, which is an important cause of invasive candidiasis in clinic. Due to the frequently use of classical antifungal agents, there are amounts of drug resistant C. albicans being isolated, causing the significantly decreasing of the efficacy of some antifungal agents in clinical treatment. Besides, the use of some compounds in clinic has been limited because of their toxicities. In such a context, drug combination therapy shows great potential on antifungal because of the synergy of different drugs or therapeutic methods that could bring, which could improve the weaknesses of single drug.
-
Key words:
- Candida albicans /
- combination therapy /
- azole /
- polyenes /
- echinocandins /
- photodynamic therapy /
- ultrasonic therapy /
- silver nanoparticles
-
转录因子ZNF24(也称KOX17或ZNF191 )是类Krüppel锌指转录因子家族的成员,N端有一SCAN结构域(也称LeR结构域),该区域不仅含有亮氨酸[1],还有选择性的异型或同型寡聚物[2];C端有四个连续的锌指模体且都是典型的类Krüppel样[2]。我们通过小鼠胚胎干细胞基因打靶,获得了ZF-12+/-(又称Zfp191,与ZNF24同源)ES细胞,并将细胞注射入小鼠的囊胚腔,得到了正常发育的ZF-12+/-小鼠,然而得到的ZF-12 −/-小鼠胚胎发育缓慢且在7.5 d左右胚胎致死[2]。最近研究表明,ZNF24通过调控微血管内皮细胞的增殖、迁移和侵袭,在内皮细胞的血管生成中起重要作用[3]。我们前期研究发现ZNF24作为一个因子拥有多种功能,比如参与激酶转录活性调控、血管增殖、大脑发育以及DNA损伤应答等[4]。
ZNF24基因最初由上海交通大学医学院的陈竺院士科研团队与复旦大学的余龙教授科研团队合作从造血细胞中克隆获得,定位于18q12.1[5]。该区域的缺失与人类多种肿瘤相关,如浸润性乳腺癌[6]、结直肠癌[7]等。余龙教授科研团队报道了ZNF24在肝癌中的不同作用:其在肝癌组织中表达上调,可通过与β-连环蛋白基因的启动子结合,激活β-连环蛋白基因转录,进而激活其下游靶基因如细胞周期蛋白 D1 (cyclin D1)基因,促进肝癌细胞的增殖[8];也可直接与DNA甲基转移酶1(DNMT1)启动子结合,激活DNMT1基因转录,引起肝癌细胞DNA甲基化改变,进而激活PI3K-AKT途径促进肝癌细胞增殖[9],提示ZNF24在肝癌中是癌基因。而在转移肝癌组织中ZNF24表达下调,ZNF24通过与DGL1(Discs Large 1) 启动子结合,激活DGL1基因转录,通过Yes相关蛋白(Yes-associated protein, YAP)信号通路抑制肝癌细胞的转移,提示ZNF24在肝癌转移中是抑癌基因[10]。此外,前列腺癌中ZNF24表达上调,通过调控Twist1促进肿瘤细胞上皮间质转换(EMT)、增殖、侵袭和转移,提示ZNF24在前列腺癌中是癌基因[11]。但是,ZNF24在甲状腺癌中表达下调,通过竞争性结合β-连环蛋白,抑制它与辅助因子LEF1/TCF1形成功能性复合物,从而抑制Wnt信号通路,进而抑制肿瘤增生与转移[12],提示ZNF24在甲状腺癌中是抑癌基因。令人感兴趣的是,研究miRNA-940(microRNA-940)在肿瘤中的作用,发现ZNF24是其调控的靶基因,在三阴乳腺癌(TNBC)中miRNA-940靶向下调ZNF24,抑制三阴乳腺癌(TNBC)细胞的增殖和转移[13],提示ZNF24促进TNBC细胞的增殖和转移是癌基因。但是,在人胃癌组织中miRNA-940 通过靶向抑制 ZNF24 表达,促进癌细胞的侵袭和转移[14],提示ZNF24抑制胃癌细胞的侵袭和转移是抑癌基因。这些结果表明,ZNF24通过调控不同的靶基因,在多种不同肿瘤的发生发展、侵袭和转移中起着重要复杂的两面性作用(促进或抑制)。
结直肠癌是一种常见的恶性肿瘤,在我国拥有较高的发病率和较低的生存率[15]。目前,结直肠癌与ZNF24的关系仍不明确。因此,我们构建ZNF24基因过表达的慢病毒载体,包装成病毒并转染结直肠癌细胞HCT116,获得了ZNF24基因过表达的HCT116细胞株,为后续研究的开展提供物质基础。
1. 实验材料
1.1 细胞及载体
293T细胞、人结直肠癌HCT116细胞、大肠杆菌感受态DH5α均来自本实验室,质粒pMT406、包装质粒pCMV-dR8.9、pCMV-VSV-G均购自上海Sangon Biotech公司。
1.2 主要试剂
限制性内切酶BamHI(R6021)、限制性内切酶XhoI(RK21100)、DNA胶回收试剂盒(AK1001)、逆转录试剂盒(RR037A)、荧光定量PCR试剂盒(RR420L)(Takara公司,日本);质粒小提试剂盒(PD1211,Promega公司,美国);无缝克隆试剂盒(C5891)、 AxyPrep 总RNA小量提取试剂盒(AP-MN-MS-RNA-250G)、兔抗ZNF24多克隆抗体(D324009)、兔抗GAPDH多克隆抗体(D110016)、山羊抗兔IgG(D111018)(Sangon Biotech公司,中国);BCA蛋白浓度测定试剂盒(P0012A)、胰酶(C0202)(碧云天生物科技公司,中国);DMEM细胞培养基(SH30022,赛默飞世尔生物科技公司,美国);胎牛血清(6170-078, Ausbian公司,澳大利亚)。
2. 实验方法
2.1 ZNF24基因过表达慢病毒载体的构建
2.1.1 目的片段获取
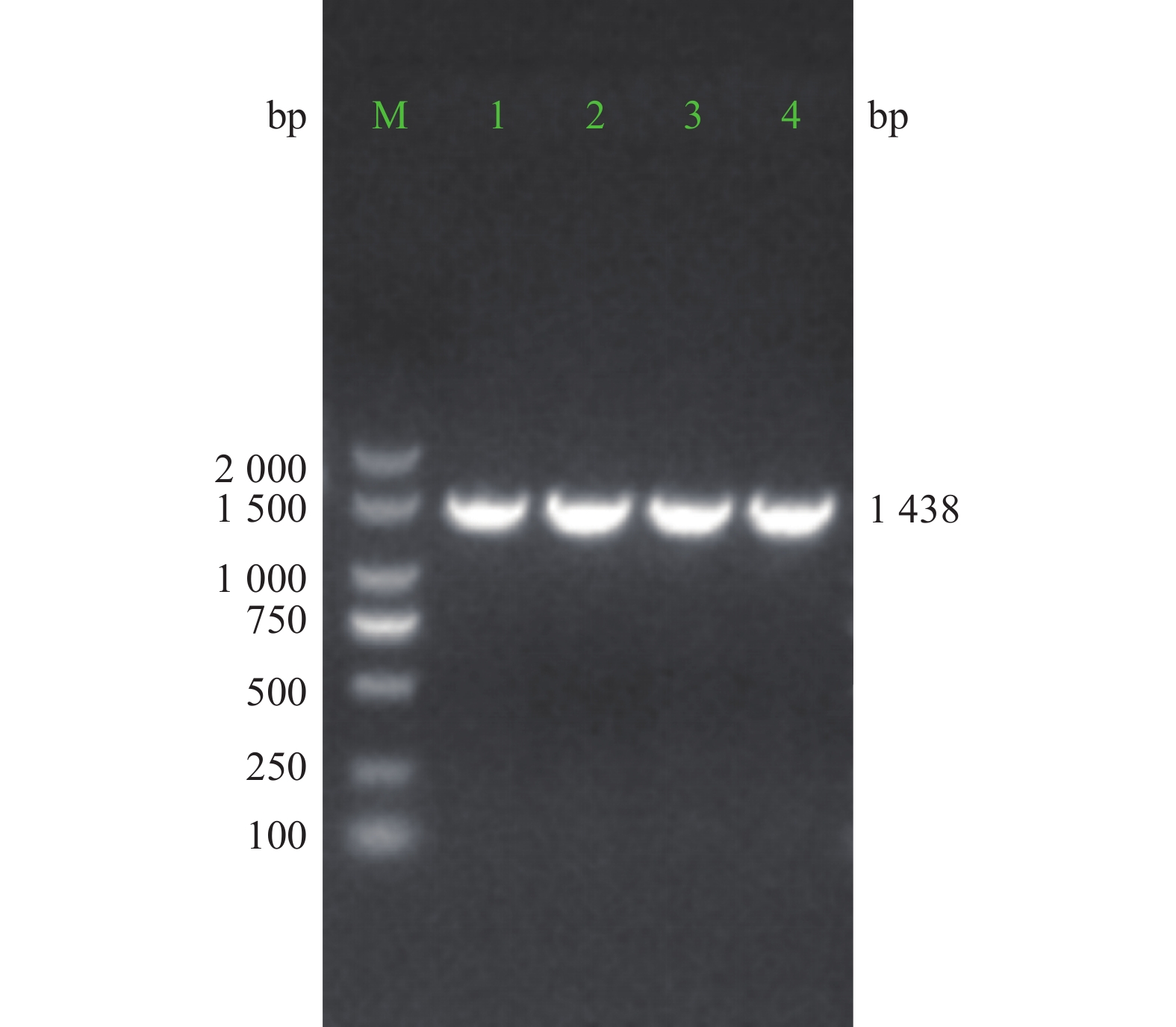
ZNF24基因、3FLAG和相关引物均由上海Sangon Biotech公司合成。Primer 1和Primer 2用于PCR扩增ZNF24,产物大小为1 128 bp;Primer 3和Primer 4用于PCR扩增3FLAG,产物大小为111 bp; Primer 5和Primer 6用于菌落PCR鉴定,阳性产物大小为1 438 bp。引物序列见表1。
表 1 ZNF24基因、3FLAG的特异性引物序列以及相关引物序列片段名称 序列 Primer 1 TGGCAAAGAATTGGATCCGCC
ACCATGTCTGCACAGTCAGTGGAAGPrimer 2 AACTTTCACAACATTCAGAAGTTTT Primer 3 CTGAATGTTGTGAAAGTTGACTACAAGGATGA Primer 4 CATAATACTAGTCTCGAGTTATTTGTCGTCATCATC Primer 5 CGGCTCTAGAGCCTCTGCTA Primer 6 CGTGAGTCAAACCGCTATCCAC ZNF24(或3FLAG)的PCR反应条件:98 ℃预变性3 min;98 ℃变性10 s,55 ℃退火15 s,72 ℃延伸1 min(或10 s),共 30个循环;72 ℃延伸10 min。
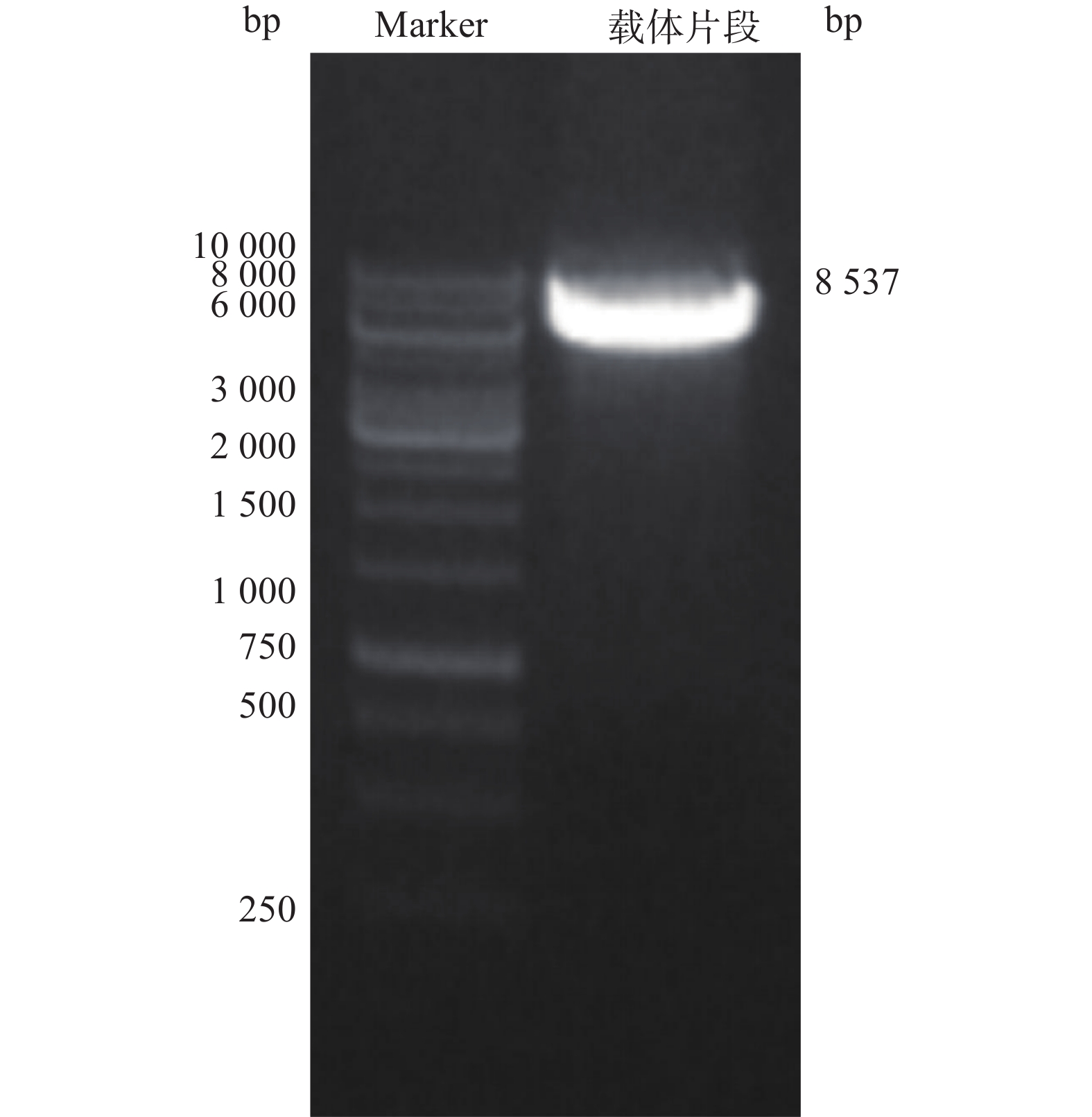
2.1.2 载体线性化、重组质粒的构建与鉴定
用限制性内切酶BamHI和XhoI酶切pMT406,胶回收线性化载体(大小约8 537 bp)。线性化的载体、PCR扩增的ZNF24与3FLAG产物,通过同源重组(无缝克隆)反应,将10 μl反应产物转化至DH5α。平皿培养过夜,挑取单克隆进行菌落PCR鉴定, 阳性克隆产物预期为1 438 bp。PCR阳性的克隆进一步测序鉴定,测序正确的重组质粒命名为pMT-ZNF24。
2.2 慢病毒载体包装、滴度测定
转染前24 h,胰酶消化并重悬293T细胞,取10个10 cm的培养皿,以1×107个/皿的细胞密度铺板。细胞贴壁后将原有培养基更换为Opti-MEM®培养基,体积为9 ml。取100 μg pMT-ZNF24(或pMT406)、65 μg pCMV-dR8.9、35 μg pCMV-VSV-G和适量Opti-MEM®培养基加入至15 ml无菌离心管中混匀,总体积为5 ml。再取500 μl细胞转染液和4.5 ml Opti-MEM®培养基混匀后滴加至上述离心管中,轻柔摇晃至均匀,室温孵育20 min。孵育完成后,将混合液分装到293T细胞中,每皿1 ml,轻轻摇晃混匀后放回培养箱。细胞培养6 h后弃上清液,加入10 ml DMEM培养基继续培养,2 d后收集细胞上清液。用60 ml 0.22 μm PVDF过滤装置过滤上清液, 4 ℃,25 000 r/min离心2 h,然后分装保存于−80 ℃冰箱。采用孔稀释法测定病毒滴度:准备5个EP管,各加入90 μl含10%FBS的高糖DMEM。EP管1中添加10 μl的待测病毒原液,EP管2中添加EP管1混合液10 μl,依次操作至EP管5。293T细胞接种到96孔板的 5个孔中,每孔约5×104个细胞,待细胞贴壁后去掉原液,依次加入EP管中的病毒液继续培养,24 h后换液,观察并记录3 d后稀释率最大孔中的荧光细胞数量。病毒滴度=荧光细胞数/病毒原液量。
2.3 慢病毒转染HCT116细胞
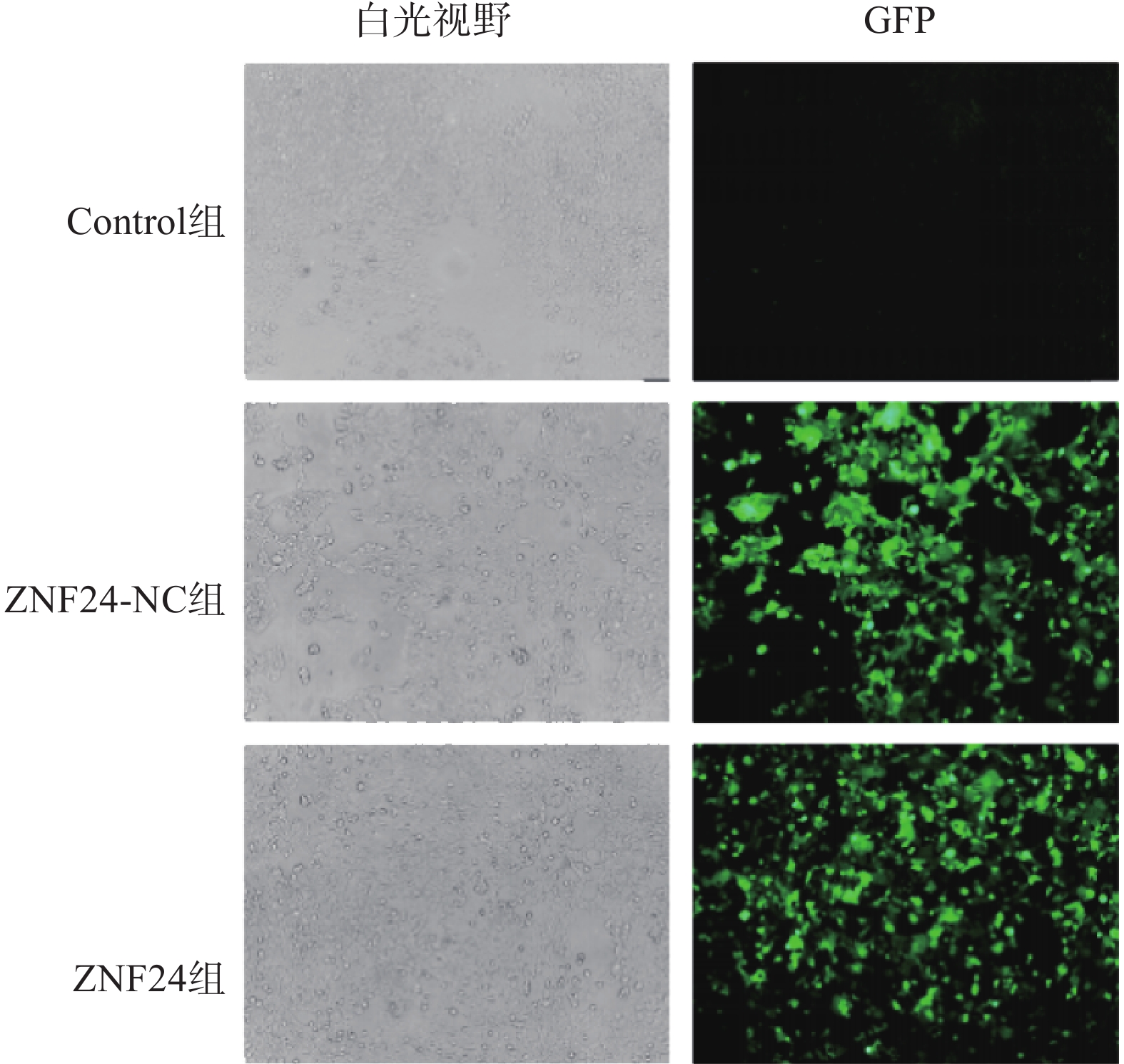
胰酶消化HCT116细胞,重悬后接种于24孔板中,每孔细胞约为3×105个,待细胞融合达30%时以细胞感染指数(MOI)=10计算病毒浓缩液体积并转染细胞。将HCT116细胞分为3组:空白对照组(HCT116细胞不转染病毒)、阴性对照组(HCT116细胞转染不含ZNF24的空载体慢病毒)和ZNF24组(HCT116细胞转染ZNF24过表达慢病毒)。转染72 h后,在ZNF24组和阴性对照组中加入 4 μg / ml嘌呤霉素,继续培养72 h后得到稳定表达细胞株。
2.4 实时荧光定量PCR检测ZNF24 mRNA的表达
慢病毒转染HCT116细胞,TRIzol法裂解细胞并提取RNA,mRNA反转录成cDNA后扩增ZNF24。ZNF24引物序列上游为CATTCCCTAAGGCACTGTGAT,下游为TTGAGGAACACCCATACTGAGA;GAPDH引物序列上游为TGACTTCAACAGCGACACCCA,下游为CACCCTGTTGCTGTAGCCAAA。2−ΔΔCt法分析ZNF24 mRNA的表达量。
2.5 蛋白印迹法检测ZNF24蛋白的表达
慢病毒转染HCT116细胞,裂解液(含蛋白酶抑制剂)裂解细胞,提取总蛋白并测定浓度。SDS-PAGE电泳后转模,脱脂牛奶封闭1 h,室温一抗孵育3 h,室温荧光素标记二抗孵育2 h,Odyssey双色红外激光成像系统检测荧光信号。
2.6 统计学分析
实验数据以3个独立试验的(
$\bar{x} \text{±} s$ )表示,采用 GraphPad Prism 5.0软件中单因素方差分析或t检验进行分析。3. 结果
3.1 PCR扩增及电泳结果
电泳结果显示,分别获得了大小约1 128 bp的ZNF24扩增产物与大小约111 bp的3FLAG扩增产物(图1)。
3.2 载体pMT406线性化
电泳结果显示,得到大小约8 537 bp线性化载体条带(图2)。
3.3 重组慢病毒载体pMT-ZNF24菌落PCR与测序鉴定
重组质粒经PCR扩增,电泳结果显示,获得约1 438 bp大小的阳性克隆PCR产物条带(图3)。对PCR产物进行测序比对分析,结果与目标序列完全一致。
3.4 病毒滴度检测结果及荧光显微镜下绿色荧光表达
ZNF24过表达慢病毒的滴度为3.25×109 TU/ml,ZNF24-NC慢病毒的滴度为6.19×109 TU/ml。以MOI=10计算病毒体积并转染HCT116细胞,4 μg /ml 嘌吟霉素筛选,荧光显微镜下约85%的细胞呈现绿色荧光蛋白(GFP)表达(图4)。
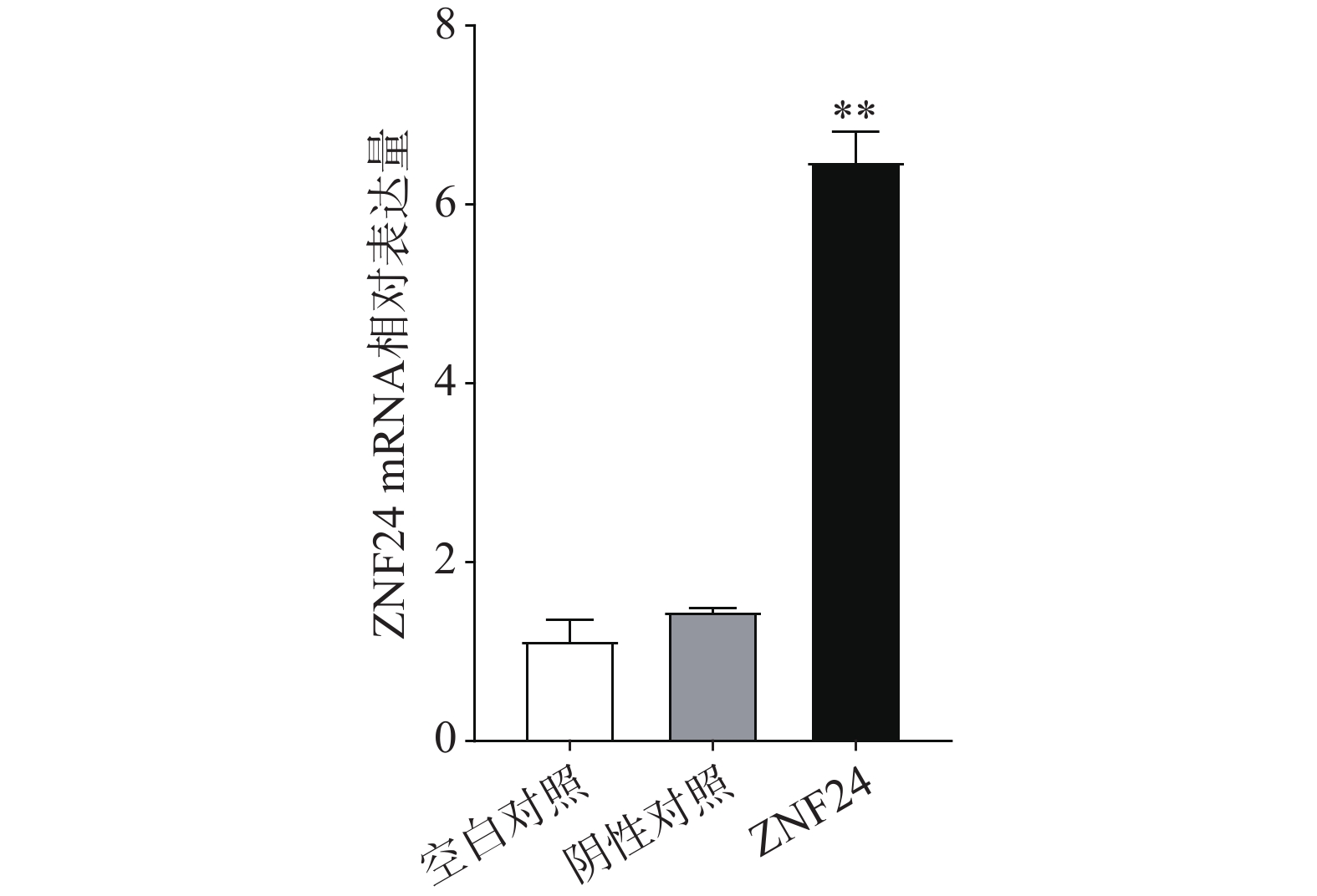
3.5 HCT116细胞中ZNF24 mRNA相对表达量比较
qRT-PCR结果表明,转染ZNF24过表达慢病毒的细胞组中ZNF24的mRNA表达显著高于阴性对照组和空白对照组(图5)。
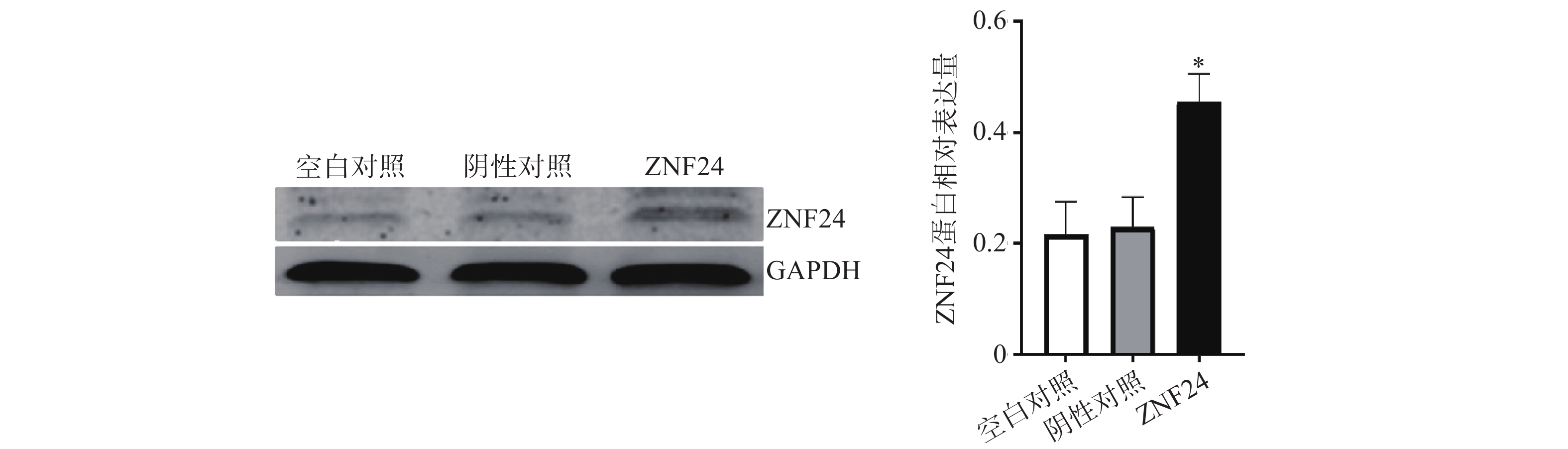
3.6 HCT116细胞中ZNF24蛋白相对表达量比较
蛋白印迹检测结果显示,转染ZNF24过表达慢病毒的细胞组中ZNF24表达水平明显高于阴性对照组和空白对照组 (图6)。
4. 讨论
结直肠癌是癌症致死的一个主要原因[16],致死的关键要素是其高水平的复发和转移[17]。结肠癌的发病机制十分复杂,涉及多种癌基因、抑癌基因的异常表达。因此,研究结直肠癌的发病机制,特别是结直肠癌复发转移的机制极其重要。研究表明,多种转录因子在结直肠癌等肿瘤中异常表达,参与结直肠癌的发病、转移和侵袭,如在肿瘤微环境参与VEGF表达调控的 STAT3转录因子已成为新的抗肿瘤药物的作用靶点[18]。
体外与乳腺癌细胞肿瘤动物模型的研究表明,ZNF24通过与血管内皮生长因子(VEGF)启动子上游序列(−144/−134, 非(TCAT) n重复序列)直接结合抑制VEGF基因转录,从而抑制血管增生达到抑制肿瘤生长[19]。然而,敲减人微血管内皮细胞中的ZNF24导致细胞迁移,侵袭和增殖减弱,暗示ZNF24具有促进人微血管内皮细胞的血管生成潜力[3]。多项研究表明ZNF24通过调控不同靶基因(如Twist1[11]、β-连环蛋白[8]和DGL1[10]等)的转录表达以及竞争性结合蛋白因子[8],在多种不同肿瘤的发生发展、侵袭和转移中起着重要复杂的两面性作用(促进或抑制)。
本研究成功构建了ZNF24过表达慢病毒载体,获得相应的病毒。转染HCT116细胞,获得稳定过表达ZNF24的HCT116细胞株,为开展后续研究提供了物质基础。
-
[1] ZENG Z R, TIAN G, DING Y H, et al. Surveillance study of the prevalence, species distribution, antifungal susceptibility, risk factors and mortality of invasive candidiasis in a tertiary teaching hospital in Southwest China[J]. BMC Infect Dis,2019,19(1):939. doi: 10.1186/s12879-019-4588-9 [2] DE CREMER K, DE BRUCKER K, STAES I, et al. Stimulation of superoxide production increases fungicidal action of miconazole against Candida albicans biofilms[J]. Sci Rep,2016,6:27463. doi: 10.1038/srep27463 [3] LOHSE M B, GULATI M, CRAIK C S, et al. Combination of antifungal drugs and protease inhibitors prevent Candida albicans biofilm formation and disrupt mature biofilms[J]. Front Microbiol,2020,11:1027. doi: 10.3389/fmicb.2020.01027 [4] ZHANG M, YAN H Y, LU M J, et al. Antifungal activity of ribavirin used alone or in combination with fluconazole against Candida albicans is mediated by reduced virulence[J]. Int J Antimicrob Agents,2020,55(1):105804. doi: 10.1016/j.ijantimicag.2019.09.008 [5] NISHIMOTO A T, SHARMA C, ROGERS P D. Molecular and genetic basis of azole antifungal resistance in the opportunistic pathogenic fungus Candida albicans[J]. J Antimicrob Chemother,2020,75(2):257-270. doi: 10.1093/jac/dkz400 [6] DIŽOVÁ S, ČERNÁKOVÁ L, BUJDÁKOVÁ H. The impact of farnesol in combination with fluconazole on Candida albicans biofilm: regulation of ERG20, ERG9, and ERG11 genes[J]. Folia Microbiol (Praha),2018,63(3):363-371. doi: 10.1007/s12223-017-0574-z [7] 周罗成, 王宁, 朱莹莹, 等. 法尼醇在促氟康唑耐药白念珠菌凋亡中的作用机制[J]. 内科理论与实践, 2020, 15(1):49-52. doi: 10.16138/j.1673-6087.2020.01.010 [8] HUANG X X, ZHENG M Y, YI Y L, et al. Inhibition of berberine hydrochloride on Candida albicans biofilm formation[J]. Biotechnol Lett,2020,42(11):2263-2269. doi: 10.1007/s10529-020-02938-6 [9] ZHANG Y, BAI X, YUWEN H S, et al. Alkaloids from Tabernaemontana divaricata combined with fluconazole to overcome fluconazole resistance in Candida albicans[J]. Bioorg Chem,2021,107:104515. doi: 10.1016/j.bioorg.2020.104515 [10] RHIMI W, ANEKE C I, ANNOSCIA G, et al. Effect of chlorogenic and Gallic acids combined with azoles on antifungal susceptibility and virulence of multidrug-resistant Candida spp. and Malassezia furfur isolates[J]. Med Mycol,2020,58(8):1091-1101. doi: 10.1093/mmy/myaa010 [11] GUO N, LING G H, LIANG X Y, et al. In vitro synergy of pseudolaric acid B and fluconazole against clinical isolates of Candida albicans[J]. Mycoses,2011,54(5):e400-e406. doi: 10.1111/j.1439-0507.2010.01935.x [12] SHIH P Y, LIAO Y T, TSENG Y K, et al. A potential antifungal effect of chitosan against Candida albicans is mediated via the inhibition of SAGA complex component expression and the subsequent alteration of cell surface integrity[J]. Front Microbiol,2019,10:602. doi: 10.3389/fmicb.2019.00602 [13] LO W H, DENG F S, CHANG C J, et al. Synergistic antifungal activity of chitosan with fluconazole against Candida albicans, Candida tropicalis, and fluconazole-resistant strains[J]. Molecules,2020,25(21):5114. doi: 10.3390/molecules25215114 [14] 严园园, 汪天明, 施高翔, 等. 黄连解毒汤联合氟康唑对耐药白念珠菌麦角甾醇的影响[J]. 中国中药杂志, 2015, 40(4):727-732. [15] SUN W W, WANG D C, YU C X, et al. Strong synergism of dexamethasone in combination with fluconazole against resistant Candida albicans mediated by inhibiting drug efflux and reducing virulence[J]. Int J Antimicrob Agents,2017,50(3):399-405. doi: 10.1016/j.ijantimicag.2017.03.015 [16] LI X Y, YU C X, HUANG X, et al. Synergistic effects and mechanisms of budesonide in combination with fluconazole against resistant Candida albicans[J]. PLoS One,2016,11(12):e0168936. doi: 10.1371/journal.pone.0168936 [17] DELARZE E, BRANDT L, TRACHSEL E, et al. Identification and characterization of mediators of fluconazole tolerance in Candida albicans[J]. Front Microbiol,2020,11:591140. doi: 10.3389/fmicb.2020.591140 [18] UPPULURI P, NETT J, HEITMAN J, et al. Synergistic effect of calcineurin inhibitors and fluconazole against Candida albicans biofilms[J]. Antimicrob Agents Chemother,2008,52(3):1127-1132. doi: 10.1128/AAC.01397-07 [19] O'MEARA T R, ROBBINS N, COWEN L E. The Hsp90 chaperone network modulates Candida virulence traits[J]. Trends Microbiol,2017,25(10):809-819. doi: 10.1016/j.tim.2017.05.003 [20] YUAN R, TU J, SHENG C Q, et al. Effects of Hsp90 inhibitor ganetespib on inhibition of azole-resistant Candida albicans[J]. Front Microbiol,2021,12:680382. doi: 10.3389/fmicb.2021.680382 [21] ZHANG J Q, LIU W, TAN J W, et al. Antifungal activity of geldanamycin alone or in combination with fluconazole against Candida species[J]. Mycopathologia,2013,175(3-4):273-279. doi: 10.1007/s11046-012-9612-1 [22] MATTHEWS R C, RIGG G, HODGETTS S, et al. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. [J]. 抗菌试剂及化学方法, 2003, 47(7): 2208-2216. MATTHEWS R C, RIGG G, HODGETTS S, et al. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90.[J]. Antimicrob Agents Chemother, 2003, 47(7):2208-2216. [23] GRELA E, ZDYBICKA-BARABAS A, PAWLIKOWSKA-PAWLEGA B, et al. Modes of the antibiotic activity of amphotericin B against Candida albicans[J]. Sci Rep,2019,9(1):17029. doi: 10.1038/s41598-019-53517-3 [24] AVERSA F, BUSCA A, CANDONI A, et al. Liposomal amphotericin B (AmBisome®) at beginning of its third decade of clinical use[J]. J Chemother,2017,29(3):131-143. doi: 10.1080/1120009X.2017.1306183 [25] KHAN S N, KHAN S, MISBA L, et al. Synergistic fungicidal activity with low doses of eugenol and amphotericin B against Candida albicans[J]. Biochem Biophys Res Commun,2019,518(3):459-464. doi: 10.1016/j.bbrc.2019.08.053 [26] CHUDZIK B, BONIO K, DABROWSKI W, et al. Synergistic antifungal interactions of amphotericin B with 4-(5-methyl-1, 3, 4-thiadiazole-2-yl) benzene-1, 3-diol[J]. Sci Rep,2019,9(1):12945. doi: 10.1038/s41598-019-49425-1 [27] UCHIDA R, KONDO A, YAGI A, et al. Simpotentin, a new potentiator of amphotericin B activity against Candida albicans, produced by Simplicillium minatense FKI-4981[J]. J Antibiot (Tokyo),2019,72(3):134-140. doi: 10.1038/s41429-018-0128-x [28] FUKUDA T, NAGAI K, YAGI A, et al. Nectriatide, a potentiator of amphotericin B activity from Nectriaceae sp. BF-0114[J]. J Nat Prod,2019,82(10):2673-2681. doi: 10.1021/acs.jnatprod.8b01056 [29] YAGI A, UCHIDA R, KOBAYASHI K, et al. Polyketide glycosides phialotides A to H, new potentiators of amphotericin B activity, produced by Pseudophialophora sp. BF-0158[J]. J Antibiot (Tokyo),2020,73(4):211-223. doi: 10.1038/s41429-019-0276-7 [30] ALVAREZ C, ANDES D R, KANG J Y, et al. Antifungal efficacy of an intravenous formulation containing monomeric amphotericin B, 5-fluorocytosine, and saline for sodium supplementation[J]. Pharm Res,2017,34(5):1115-1124. doi: 10.1007/s11095-017-2121-7 [31] PERLIN D S. Mechanisms of echinocandin antifungal drug resistance[J]. Ann N Y Acad Sci,2015,1354(1):1-11. doi: 10.1111/nyas.12831 [32] LARWOOD D J. Nikkomycin Z—ready to meet the promise? JoF,2020,6(4):261. doi: 10.3390/jof6040261 [33] KOVÁCS R, NAGY F, TÓTH Z, et al. Synergistic effect of nikkomycin Z with caspofungin and micafungin against Candida albicans and Candida parapsilosis biofilms[J]. Lett Appl Microbiol,2019,69(4):271-278. doi: 10.1111/lam.13204 [34] CHEN Y L, LEHMAN V N, AVERETTE A F, et al. Posaconazole exhibits in vitro and in vivo synergistic antifungal activity with caspofungin or FK506 against Candida albicans[J]. PLoS One,2013,8(3):e57672. doi: 10.1371/journal.pone.0057672 [35] ROBBINS N, SPITZER M, YU T, et al. An antifungal combination matrix identifies a rich pool of adjuvant molecules that enhance drug activity against diverse fungal pathogens[J]. Cell Rep,2015,13(7):1481-1492. doi: 10.1016/j.celrep.2015.10.018 [36] COOLS T L, STRUYFS C, DRIJFHOUT J W, et al. A linear 19-mer plant defensin-derived peptide acts synergistically with caspofungin against Candida albicans biofilms[J]. Front Microbiol,2017,8:2051. doi: 10.3389/fmicb.2017.02051 [37] TROSKIE A M, RAUTENBACH M, DELATTIN N, et al. Synergistic activity of the tyrocidines, antimicrobial cyclodecapeptides from Bacillus aneurinolyticus, with amphotericin B and caspofungin against Candida albicans biofilms[J]. Antimicrob Agents Chemother,2014,58(7):3697-3707. doi: 10.1128/AAC.02381-14 [38] MASOUDI Y, VAN RENSBURG W, BARNARD-JENKINS B, et al. The influence of cellulose-type formulants on anti- Candida activity of the tyrocidines[J]. Antibiotics (Basel),2021,10(5):597. doi: 10.3390/antibiotics10050597 [39] SABINO C P, WAINWRIGHT M, RIBEIRO M S, et al. Global priority multidrug-resistant pathogens do not resist photodynamic therapy[J]. J Photochem Photobiol B,2020,208:111893. doi: 10.1016/j.jphotobiol.2020.111893 [40] HU X Q, HUANG Y Y, WANG Y G, et al. Antimicrobial photodynamic therapy to control clinically relevant biofilm infections[J]. Front Microbiol,2018,9:1299. doi: 10.3389/fmicb.2018.01299 [41] PANARIELLO B H D, KLEIN M I, ALVES F, et al. DNase increases the efficacy of antimicrobial photodynamic therapy on Candida albicans biofilms[J]. Photodiagnosis Photodyn Ther,2019,27:124-131. doi: 10.1016/j.pdpdt.2019.05.038 [42] DAVIES A, GEBREMEDHIN S, YEE M, et al. Cationic porphyrin-mediated photodynamic inactivation of Candida biofilms and the effect of miconazole[J]. J Physiol Pharmacol,2016,67(5):777-783. [43] LU J J, LI W, ZHENG W A, et al. Successful treatment of kerion with itraconazole and ALA-PDT: a case report[J]. Photodiagnosis Photodyn Ther,2019,27:385-387. doi: 10.1016/j.pdpdt.2019.07.007 [44] YANG M, DU K Y, HOU Y R, et al. Synergistic antifungal effect of amphotericin B-loaded poly(lactic-co-glycolic acid) nanoparticles and ultrasound against Candida albicans biofilms[J]. Antimicrob Agents Chemother,2019,63(4):e02022-e02018. [45] Gong-chang YU, Yong ZHANG, Ke NIE. Anti-emetic mechanisms of Xiaobanxia Tang Decoction on the chemotherapy-induced pica model in rats[J]. 中国药理学与毒理学杂志, 2015, 29(S1): 84-85. Gong-chang YU, Yong ZHANG, Ke NIE. Anti-emetic mechanisms of Xiaobanxia Tang Decoction on the chemotherapy-induced pica model in rats[J]. Chinese Journal of Pharmacology and Toxicology, 2015, 29(S1):84-85. [46] RADHAKRISHNAN V S, REDDY MUDIAM M K, KUMAR M, et al. Silver nanoparticles induced alterations in multiple cellular targets, which are critical for drug susceptibilities and pathogenicity in fungal pathogen (Candida albicans)[J]. Int J Nanomedicine,2018,13:2647-2663. doi: 10.2147/IJN.S150648 [47] LARA H H, LOPEZ-RIBOT J L. Inhibition of mixed biofilms of Candida albicans and methicillin-resistant Staphylococcus aureus by positively charged silver nanoparticles and functionalized silicone elastomers[J]. Pathogens,2020,9(10):784. doi: 10.3390/pathogens9100784 [48] GUERRERO D J P, BONILLA J J A, LÓPEZ C C O, et al. Encapsulation of silver nanoparticles in polylactic acid or poly(lactic-co-glycolic acid) and their antimicrobial and cytotoxic activities[J]. J Nanosci Nanotechnol,2019,19(11):6933-6941. doi: 10.1166/jnn.2019.16663 [49] LEE B, LEE M J, YUN S J, et al. Silver nanoparticles induce reactive oxygen species-mediated cell cycle delay and synergistic cytotoxicity with 3-bromopyruvate in Candida albicans, but not in Saccharomyces cerevisiae[J]. Int J Nanomedicine,2019,14:4801-4816. doi: 10.2147/IJN.S205736 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 7517
- HTML全文浏览量: 2358
- PDF下载量: 50
- 被引次数: 0
 下载:
下载:
