

 下载:
下载:


 下载:
下载: 


-
系统性红斑狼疮(systemic lupus erythematosus, SLE)是一种慢性自身免疫性疾病,在我国的发病率为40~70/10万[1],是导致女性死亡的十大原因之一。SLE发病机制不明,尚无有效治疗手段。目前主要以激素、环磷酰胺和骁悉等免疫抑制剂为主,但药物引起的股骨头坏死、重症感染等副作用严重影响治疗效果[2]。随着对其免疫病理机制研究的进展,针对免疫紊乱关键环节的调控策略,如生物制剂、血浆置换、干细胞等为SLE的治疗提供了新的视角和理念[3-6]。但是,这些治疗方法仍然存在很多局限性和个体差异。SLE在祖国医学中属“阴阳毒” “蝴蝶丹” “痈证”等。近年来,很多中医药单独或者联合西药治疗SLE取得较好的效果,且副作用少,价格低廉,逐渐受到广大临床医生的认可和患者的接受。但是中医药治疗SLE的确切调节机制和作用环节尚不明确,本文从代谢组学、免疫细胞、淋巴细胞因子和细胞凋亡等多个方面,对中医药治疗SLE机制的研究进行综述,为利用现代化方法探索祖国传统医学治疗SLE的机制研究提供思路。
-
SLE是一种多器官和组织受累的自身免疫性疾病,临床表现多样,目前在现代医学领域尚无疾病亚型的分类。1987年中国卫生部药政局颁布的《系统性红斑狼疮辨证标准》,将SLE辨证分为4种证候,即热毒炽盛、肝肾阴虚、气血瘀滞、脾肾阳虚;1993年《中药新药治疗红斑狼疮的临床研究指导原则》将SLE分为热毒炽热、阴虚内热、肝肾阴虚、瘀热伤肝、脾肾阳虚、风湿热痹6种证候。
SLE的临床表现众多,但是其本质均是肝肾阴虚,进而产生热毒和血瘀。首先肝肾阴虚容易导致患者成为SLE易感人群,热毒和血瘀则可导致多器官和组织的损伤,因此,临床最常见的SLE证候为热毒血瘀阴虚证[7]。此外,中医证型与SLE不同系统受累存在关联,其中热毒炽盛证与神经精神系统、脾肾阳虚证与肾脏系统、瘀热痹阻证与血液和心血管系统、阴虚内热与肺脏系统存在相关性。有人利用气相色谱-质谱联用技术,对不同SLE中医证候类型患者的尿液进行了分析,发现相对于正常对照组,阴虚内热和脾肾阳虚两种证型的SLE患者有4种相同的代谢物(1,2,3-三羧酸环丙烷、柠檬酸三乙酯、甘氨酸和氯苯甲酸);脾肾阳虚和阴虚内热两种证型的SLE患者有特征代谢物(包括琥珀酸、马尿酸、赤霉酸、9,10-开环胆甾-5,7,10-三烯和2H苯并吡喃),提示三羧酸循环、肠道菌群和糖代谢等通路参与SLE不同证型的病理机制[8]。
体外实验发现,热毒血瘀型SLE患者来源的血清在体外刺激人肾系膜细胞可诱导特异蛋白表达,其中膜联蛋白A2和 A5可作为热毒血瘀证的生物标志物及潜在治疗靶点[9]。自身抗体是SLE诊断的重要指标之一,基于不同自身抗体种类和SLE常见中医证型的关联研究发现:抗核抗体滴度按均值大小排列,依次为热毒炽盛型、阴虚内热型和脾肾阳虚型;抗双链DNA抗体(dsDNA)滴度按阳性率大小排列,依次是脾肾阳虚型、阴虚内热型和热毒炽盛型,其中抗组蛋白抗体阳性率在脾肾阳虚型和热毒炽盛型中存在显著差异[10]。还有研究发现SLE患者的自身抗体谱可以指导SLE中医辨证分型,比如,抗dsDNA抗体和抗U1RNP抗体阳性,多考虑为风湿热痹型,而抗dsDNA抗体和抗核小体抗体阳性,多考虑为脾肾阳虚型[11]。另外,SLE患者的血瘀证与抗心磷脂抗体、补体C3和补体C4等免疫学指标存在相关性,提示自身免疫复合物形成可能是血瘀证的发病机理之一[11]。
-
SLE是一种多器官受累的系统性疾病,患者体内的代谢网络必然发生变化,因此,患者循环系统中代谢物种类、浓度或相对比例都有可能发生改变。代谢组学是一种系统生物学技术,通过对体液及组织中的小分子代谢物的整体组成进行动态的跟踪监测,可以分析机体在环境和病理刺激时内源代谢物的改变,为临床疾病生物标志物探索、疾病分型和疗效评估等研究提供靶标和线索。多数研究显示SLE患者与健康对照组相比,血清代谢物的差异主要集中在脂质和氨基酸。高密度脂蛋白(HDL)降低,低密度脂蛋白(LDL)和极低密度脂蛋白(VLDL)升高,均提示SLE患者存在脂质代谢紊乱,而血清氨基酸水平降低和糖酵解中间代谢产物丙酮酸下降则说明SLE患者存在能量代谢紊乱[12]。
解毒祛瘀滋阴方能有效控制SLE的病情进展,研究报道其临床效果显著优于单用糖皮质激素组。在代谢水平上,该中医复方可显著上调二十碳五烯酸的含量,同时显著上调氨基酸生物标志物甘氨酸、苯丙氨酸和色氨酸至正常水平 [13]; 在狼疮小鼠模型的研究中,昆仙胶囊治疗组与对照组比较,其小鼠血清中支链氨基酸、丙氨酸、糖蛋白、 柠檬酸、磷酸胆碱、牛磺酸和葡萄糖含量升高,而酮体、肌酸等含量降低,提示其发挥作用的机制可能与促进机体正常的能量代谢和氧化能力,稳定细胞膜以及抑制细胞的异常凋亡有关[14]。MRL/lps狼疮鼠存在较明显的脂质代谢异常,青蒿-鳖甲药可以有效改善狼疮鼠体内脂质代谢异常,降低血清L-a-溶血卵磷脂水平,减少血栓发生,同时,改善狼疮小鼠急性期的炎症反应,减少其病理损伤[15]。
-
巨噬细胞是机体固有免疫的重要组成细胞,能够通过吞噬凋亡细胞协助清除自身抗原,避免自身免疫反应的发生。巨噬细胞吞噬能力障碍在SLE发病过程中发挥重要作用。体外研究显示植物多糖、柴胡多糖和黑灵芝多糖都能够显著增强巨噬细胞的吞噬能力[16],可能通过调控固有免疫应答影响SLE的病程。雷公藤多苷能够下调SLE患者树突细胞表面的Toll样受体9(TLR9),进而抑制树突细胞对T细胞的激活作用[17]。
淋巴细胞分化异常和过度激活是SLE的主要病理机制之一,包括T、B淋巴细胞过度活化和炎性细胞因子的异常表达。白藜芦醇能够通过减少干扰素-γ等细胞因子,抑制CD4+T细胞向Th1细胞分化,减轻降植烷(pristane)诱导狼疮鼠的炎症反应[18]。淫羊藿素能够通过转录因子STAT5b调控Foxp3/IL-17α的表达,影响调节性T(Treg)细胞和T辅助17(Th17)细胞的平衡,减轻MRL/lpr狼疮鼠的肾损伤[19]。狼疮颗粒具有补肾化毒的作用。研究发现,和西药组相比,联合狼疮颗粒的中西医结合治疗组的临床疗效更优,能够显著下调血清可溶白细胞介素2受体(sIL-2R)和肿瘤坏死因子α(TNF-α)的水平[20]。CD229即淋巴细胞信号活化分子家族3(SLAMF3),是T细胞活化和分化的重要调节分子,能够通过上调CD25,促进CD4+T细胞向具有免疫抑制功能的Treg细胞转化。在MRL/lpr狼疮鼠的研究中也发现解毒祛瘀滋肾方能够通过CD299/CD25轴促进Treg细胞分化来改善SLE病情[21]。另外,研究显示白芍总苷单药能够有效降低小鼠尿蛋白水平,同时降低其外周血单个核细胞中IL-17的mRNA水平,提示白芍总苷也能调控Th17/Treg细胞平衡这一机制发挥作用[22]。基于流式细胞术的分析,进一步证实了白芍总苷能够显著降低MRL/lpr 狼疮鼠外周血Th17细胞的比例,同时增高Treg 细胞的比例[23]。B细胞异常激活产生大量致病性自身抗体是SLE的另一重要免疫病理机制。青蒿素具有免疫调控作用,有望成为SLE的新型治疗药物,目前已进入三期临床试验[24]。在MRL/lpr 狼疮鼠的研究中发现,青蒿素衍生物马来酸蒿薏米胺(SM934)能够显著抑制B细胞TLR7/9的表达,抑制B细胞活化和浆细胞形成,从而降低自身抗体的水平,使狼疮疾病得到有效控制[25]。
-
目前,多数研究证实炎性因子的异常表达参与SLE的发生和发展。SLE患者外周血中IL-2水平较低,但IL-4、IL-6、IL-8、IL-10、IL-17、IL-21、TNF和IFN 水平较高。狼疮定具有滋阴、解毒的祛瘀作用。临床研究发现SLE 患者的sIL-2R明显高于健康对照组,且疾病活动度高的患者其表达水平更高,这可能是导致SLE患者血清中IL-2水平较低的原因,狼疮定能够有效降低SLE患者sIL-2R水平,调控细胞免疫功能,使紊乱的免疫内环境趋于平衡[26]。狼疮方具有解毒活血的功效,在慢性移植物抗宿主的小鼠模型研究中发现,狼疮方能够有效减少2型T辅助(Th2)细胞分泌的IL-6和IL-10的水平,同时降低自身抗体水平,从而减轻狼疮肾炎的损伤[27]。中药组方通过养阴清热、凉血活血等功效,有效降低阴虚内热型SLE患者血清中IL-4和IL-10水平,同时增加IL-2水平,以调控Th1/Th2细胞平衡并降低狼疮疾病的活动度[28]。凉血化瘀方能有效降低MRL/lpr狼疮鼠血清中IL-6、IL-17、IL-21和自身抗体的水平[29]。益气养阴方通过活血通络、抗炎消肿、解毒清热等功效,显著降低SLE 患者血清中IL-6、IL-8和TNF-α的水平,有助于SLE患者免疫功能的重建[30]。另有研究发现,与常规西药治疗组相比,联合滋阴降火方(生地黄、赤芍、丹参、防风、百合、青蒿、黄芪)治疗阴虚内热型SLE患者,能够显著降低血清中IL-10 和IL-18的水平并提高临床疗效,不良反应无显著差异[31]。
细胞黏附分子(ICAMs)是一类能够介导免疫细胞与内皮细胞相互作用的糖蛋白分子。研究显示ICAM-1、ICAM-3和p选择素在SLE患者血清中显著高于正常对照组,养阴清热煎通过滋阴凉血、解毒化瘀、补虚泻实等功效,能够显著抑制SLE患者高表达的ICAMs,实现对器官损伤的保护和修复[32]。血管炎是SLE的基本病理机制,血管内皮细胞是直接受损的靶器官。可溶性血栓调节蛋白 、血管假性血友病因子均是由血管内皮细胞合成的糖蛋白,也是血管内皮细胞受损的标志。在自发性狼疮小鼠模型的研究中发现,清养透解方(青蒿、鳖甲、水牛角、生地黄、牡丹皮、玄参)能够有效降低狼疮小鼠血清可溶性血栓调节蛋白、血管假性血友病因子的表达,保护血管内皮,改善肾脏组织慢性炎症的病理改变[33]。
-
活动期SLE患者外周淋巴细胞的凋亡水平显著增高,可能与Fas的高表达有关。芪加真武汤可以通过干扰Fas的表达抑制异常的细胞凋亡,从而实现治疗SLE的功效[34]。采用清热化瘀滋阴方,联合常规激素治疗气阴两虚型SLE患者,相对于常规激素单独治疗组,其外周血单个核细胞的凋亡指标bcl-2、bax、fas、p53和c-myc显著降低,提示清热化瘀滋阴方可通过降低细胞凋亡率来实现对免疫功能的调控[35]。解毒祛瘀滋阴方联合激素治疗相对单用激素治疗能够有效上调CD4和CD8阳性细胞中bcl-2的表达,促使机体紊乱的免疫内环境趋于平衡[36]。在BXSB狼疮小鼠的研究中发现,狼疮静能够显著抑制小鼠脾脏中T、B淋巴细胞的异常凋亡,阻止过量核酸抗原的释放,从而抑制B细胞产生致病性自身抗体,来达到治疗SLE的目的[37]。
细胞焦亡(pyroptosis)是一种新发现的程序性细胞死亡。核苷酸结合寡聚化结构域样受体蛋白 3 (NL- RP3)、半胱氨酸天冬氨酸蛋白酶 1 (Caspase-1)及 IL-1β 是参与细胞焦亡的重要因子,提示细胞焦亡与炎症反应的发生发展密切相关。滋阴清热方具有滋阴清热、解毒凉血的功效,联合糖皮质激素治疗阴虚内热型SLE患者效果显著,通过狼疮鼠的体内实验研究发现,滋阴清热方能够显著降低细胞焦亡相关因子,可能是其治疗SLE的机制之一[38]。
综上所述,很多中药能够通过多靶点调控机体的免疫和代谢功能,从而实现对SLE病情的改善(表1)。
表 1 中药治疗系统性红斑狼疮的机制和靶点
药名 组成成分 作用机制/靶点 相关分子 单药 柴胡多糖 巨噬细胞 NO、TNF-α、IL-1β、IL-6 黑灵芝多糖 巨噬细胞 TLR4 雷公藤素 树突细胞 TLR9 淫羊藿 Treg/Th17 STAT5b 白芍总苷 Treg/Th17 IL-17 mRNA 青蒿素 B细胞 TLR7/9 复方 青蒿-鳖甲药 青蒿、鳖甲 脂质代谢 L-a-溶血卵磷脂 解毒祛瘀滋阴方 白花蛇草、紫草、赤芍、生地、炙鳖甲 氨基酸代谢 二十碳五烯酸、甘氨酸、
苯丙氨酸、色氨酸细胞凋亡 Bcl-2 清热化瘀滋阴方 麦冬、赤芍、水牛角片、炒白术、连翘、牡丹皮、
紫草、秦艽、青风藤、金银花、白花蛇草细胞凋亡 Bcl-2、bax、fas、p53、c-myc 解毒祛瘀滋肾方 生地黄、炙鳖甲、青蒿、白花蛇草、积雪草、赤芍、
牡丹皮、佛手、升麻、重楼、生甘草Treg CD299、CD25 凉血化瘀方 水牛角、生地黄、赤芍、牡丹皮、制大黄、玄参、知母 细胞因子 IL-6、IL-17、IL-21 益气养阴方 生薏仁、丹皮、芍药、白花蛇草、升麻、水牛角、丹参、青蒿、鳖甲 细胞因子 IL-6、IL-8、TNF-α 养阴清热煎 生地黄、牡丹皮、青蒿、白花蛇草、蛇莓、半枝莲、益母草、丹参 细胞黏附分子 ICAM-1、ICAM-3 滋阴清热方 山茱萸、生地黄、茯苓、泽泻、牡丹皮、青蒿、甘草 细胞凋亡 IL-1β、NLRP3 芪加真武汤 茯苓、白芍、附子、生姜、甘草、黄芪、刺五加 细胞凋亡 fas 中药组方 生地、黄芩、积雪草、川穹、丹参、猫抓草 细胞因子 IL-2、IL-4、IL-10 狼疮静 生地、熟地、丹皮、白花蛇草 细胞凋亡、B细胞 核酸抗原 狼疮定 白花蛇草、赤芍、生地、水牛角等 细胞因子 IL-2 狼疮方 白花蛇草、半枝莲、紫草、丹参、益母草 细胞因子 IL-6、IL-10 狼疮颗粒 生地黄、熟地黄、牡丹皮、白花蛇草 Treg/Th17 sIL-2R、TNF-α 昆仙胶囊 昆明山海棠、淫羊藿、枸杞子、菟丝子等 能量代谢 支链氨基酸、丙氨酸、糖蛋白、柠檬酸、磷酸胆碱、牛磺酸、葡萄糖 注:NLRP. 核苷酸结合寡聚化结构域样受体蛋白;IL. 白介素;ICAM. 细胞黏附分子;Treg. 调节性T细胞;Th17. T辅助细胞17;TNF. 肿瘤坏死因子;TLR. Toll 样受体 -
SLE是一种复杂的多器官受累疾病,具有明显的异质性,而中医药的优势在于整体调控和辨证论治。中医理论中的“扶正驱邪”,即达到驱邪而不伤正,有助免疫功能恢复新的平衡;“同病异治”,即同一种疾病根据不同的证型采用不同的治疗方法,有助SLE个体化治疗方案的制定。但目前中医药治疗SLE的基础理论尚不明确,基于生物标志物探索中医药治疗SLE的病理机制,有望为SLE的诊疗提供新的思路和策略。
Research progress of tradition Chinese medicine treatment mechanism of systemic lupus erythematosus based on biomarkers
-
摘要: 系统性红斑狼疮(SLE)是一种多器官受累的自身免疫性疾病,目前基于糖皮质激素和免疫抑制剂的治疗,仍然存在很多局限性和个体差异。近年来,越来越多的研究表明,联合中医药治疗SLE具有疗效好,不良反应低和安全性高等优点。但是,中医药治疗SLE的确切调节机制和作用环节尚不明确,本文从代谢组学、免疫细胞、淋巴细胞因子和细胞凋亡等,对中医药治疗SLE机制的研究进行综述,为利用现代化方法探索祖国传统医学治疗SLE的机制研究提供思路。Abstract: Systemic lupus erythematosus (SLE) is an autoimmune disease with multiple organ involvement. There are still many limitations and individual differences in the treatment based on glucocorticoids and immunosuppressants. In recent years, more and more studies have shown that the combination of traditional Chinese medicine in the treatment of SLE has the advantages of good efficacy, low adverse reactions, and high safety. However, the exact regulatory mechanism and combined traditional Chinese medicine in the treatment of SLE are still unclear. This paper reviews the research on the mechanism of traditional Chinese medicine in the treatment of SLE from metabonomic, immune cells, lymphocyte factors and apoptosis, etc, provides ideas for exploring the mechanism of traditional Chinese medicine in the treatment of SLE with modern methods.
-
Key words:
- biomarkers /
- systemic lupus erythematosus (SLE) /
- traditional Chinese medicine /
- metabonomic /
- immune cells /
- cytokines
-
转录因子ZNF24(也称KOX17或ZNF191 )是类Krüppel锌指转录因子家族的成员,N端有一SCAN结构域(也称LeR结构域),该区域不仅含有亮氨酸[1],还有选择性的异型或同型寡聚物[2];C端有四个连续的锌指模体且都是典型的类Krüppel样[2]。我们通过小鼠胚胎干细胞基因打靶,获得了ZF-12+/-(又称Zfp191,与ZNF24同源)ES细胞,并将细胞注射入小鼠的囊胚腔,得到了正常发育的ZF-12+/-小鼠,然而得到的ZF-12 −/-小鼠胚胎发育缓慢且在7.5 d左右胚胎致死[2]。最近研究表明,ZNF24通过调控微血管内皮细胞的增殖、迁移和侵袭,在内皮细胞的血管生成中起重要作用[3]。我们前期研究发现ZNF24作为一个因子拥有多种功能,比如参与激酶转录活性调控、血管增殖、大脑发育以及DNA损伤应答等[4]。
ZNF24基因最初由上海交通大学医学院的陈竺院士科研团队与复旦大学的余龙教授科研团队合作从造血细胞中克隆获得,定位于18q12.1[5]。该区域的缺失与人类多种肿瘤相关,如浸润性乳腺癌[6]、结直肠癌[7]等。余龙教授科研团队报道了ZNF24在肝癌中的不同作用:其在肝癌组织中表达上调,可通过与β-连环蛋白基因的启动子结合,激活β-连环蛋白基因转录,进而激活其下游靶基因如细胞周期蛋白 D1 (cyclin D1)基因,促进肝癌细胞的增殖[8];也可直接与DNA甲基转移酶1(DNMT1)启动子结合,激活DNMT1基因转录,引起肝癌细胞DNA甲基化改变,进而激活PI3K-AKT途径促进肝癌细胞增殖[9],提示ZNF24在肝癌中是癌基因。而在转移肝癌组织中ZNF24表达下调,ZNF24通过与DGL1(Discs Large 1) 启动子结合,激活DGL1基因转录,通过Yes相关蛋白(Yes-associated protein, YAP)信号通路抑制肝癌细胞的转移,提示ZNF24在肝癌转移中是抑癌基因[10]。此外,前列腺癌中ZNF24表达上调,通过调控Twist1促进肿瘤细胞上皮间质转换(EMT)、增殖、侵袭和转移,提示ZNF24在前列腺癌中是癌基因[11]。但是,ZNF24在甲状腺癌中表达下调,通过竞争性结合β-连环蛋白,抑制它与辅助因子LEF1/TCF1形成功能性复合物,从而抑制Wnt信号通路,进而抑制肿瘤增生与转移[12],提示ZNF24在甲状腺癌中是抑癌基因。令人感兴趣的是,研究miRNA-940(microRNA-940)在肿瘤中的作用,发现ZNF24是其调控的靶基因,在三阴乳腺癌(TNBC)中miRNA-940靶向下调ZNF24,抑制三阴乳腺癌(TNBC)细胞的增殖和转移[13],提示ZNF24促进TNBC细胞的增殖和转移是癌基因。但是,在人胃癌组织中miRNA-940 通过靶向抑制 ZNF24 表达,促进癌细胞的侵袭和转移[14],提示ZNF24抑制胃癌细胞的侵袭和转移是抑癌基因。这些结果表明,ZNF24通过调控不同的靶基因,在多种不同肿瘤的发生发展、侵袭和转移中起着重要复杂的两面性作用(促进或抑制)。
结直肠癌是一种常见的恶性肿瘤,在我国拥有较高的发病率和较低的生存率[15]。目前,结直肠癌与ZNF24的关系仍不明确。因此,我们构建ZNF24基因过表达的慢病毒载体,包装成病毒并转染结直肠癌细胞HCT116,获得了ZNF24基因过表达的HCT116细胞株,为后续研究的开展提供物质基础。
1. 实验材料
1.1 细胞及载体
293T细胞、人结直肠癌HCT116细胞、大肠杆菌感受态DH5α均来自本实验室,质粒pMT406、包装质粒pCMV-dR8.9、pCMV-VSV-G均购自上海Sangon Biotech公司。
1.2 主要试剂
限制性内切酶BamHI(R6021)、限制性内切酶XhoI(RK21100)、DNA胶回收试剂盒(AK1001)、逆转录试剂盒(RR037A)、荧光定量PCR试剂盒(RR420L)(Takara公司,日本);质粒小提试剂盒(PD1211,Promega公司,美国);无缝克隆试剂盒(C5891)、 AxyPrep 总RNA小量提取试剂盒(AP-MN-MS-RNA-250G)、兔抗ZNF24多克隆抗体(D324009)、兔抗GAPDH多克隆抗体(D110016)、山羊抗兔IgG(D111018)(Sangon Biotech公司,中国);BCA蛋白浓度测定试剂盒(P0012A)、胰酶(C0202)(碧云天生物科技公司,中国);DMEM细胞培养基(SH30022,赛默飞世尔生物科技公司,美国);胎牛血清(6170-078, Ausbian公司,澳大利亚)。
2. 实验方法
2.1 ZNF24基因过表达慢病毒载体的构建
2.1.1 目的片段获取
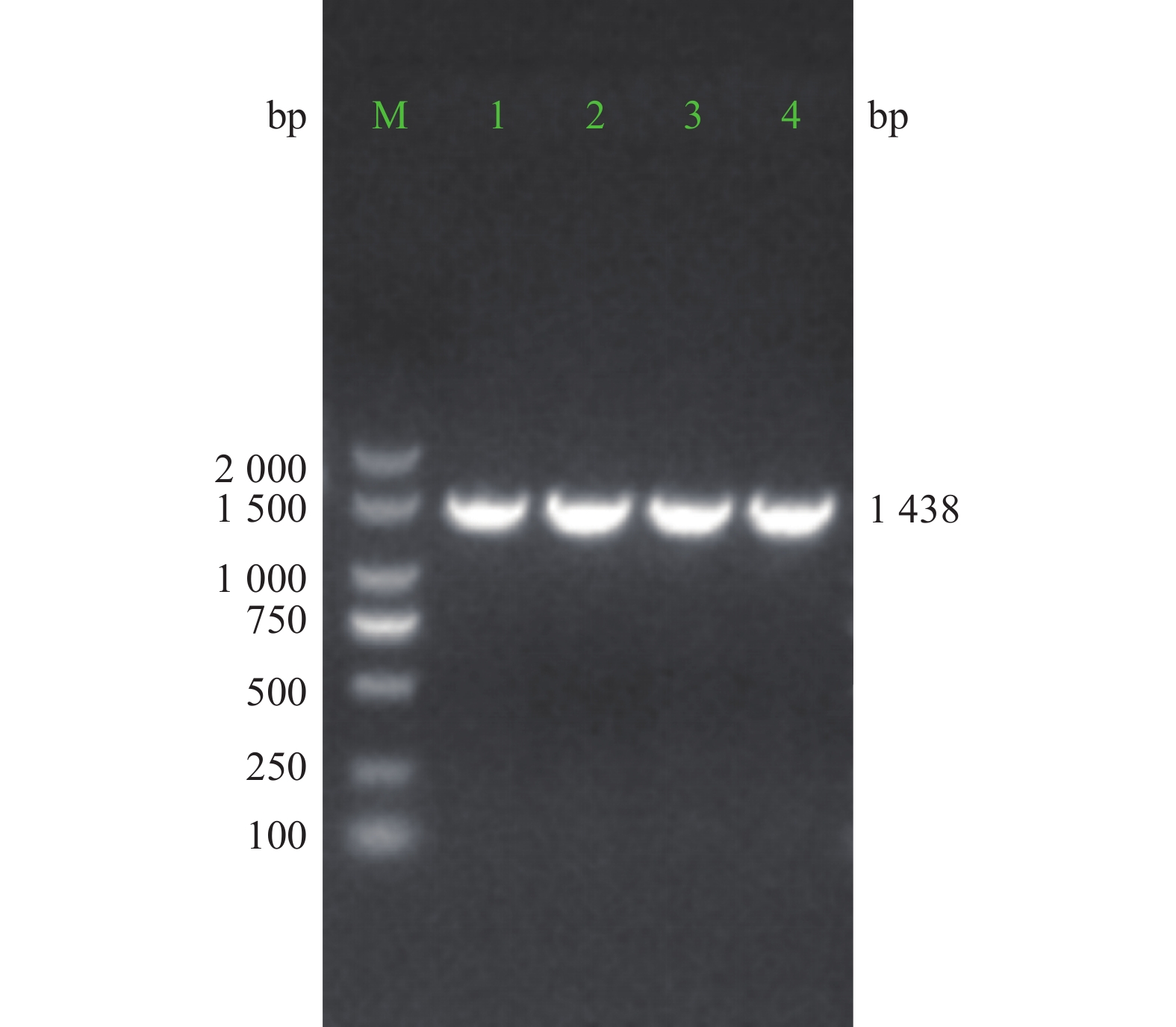
ZNF24基因、3FLAG和相关引物均由上海Sangon Biotech公司合成。Primer 1和Primer 2用于PCR扩增ZNF24,产物大小为1 128 bp;Primer 3和Primer 4用于PCR扩增3FLAG,产物大小为111 bp; Primer 5和Primer 6用于菌落PCR鉴定,阳性产物大小为1 438 bp。引物序列见表1。
表 1 ZNF24基因、3FLAG的特异性引物序列以及相关引物序列片段名称 序列 Primer 1 TGGCAAAGAATTGGATCCGCC
ACCATGTCTGCACAGTCAGTGGAAGPrimer 2 AACTTTCACAACATTCAGAAGTTTT Primer 3 CTGAATGTTGTGAAAGTTGACTACAAGGATGA Primer 4 CATAATACTAGTCTCGAGTTATTTGTCGTCATCATC Primer 5 CGGCTCTAGAGCCTCTGCTA Primer 6 CGTGAGTCAAACCGCTATCCAC ZNF24(或3FLAG)的PCR反应条件:98 ℃预变性3 min;98 ℃变性10 s,55 ℃退火15 s,72 ℃延伸1 min(或10 s),共 30个循环;72 ℃延伸10 min。
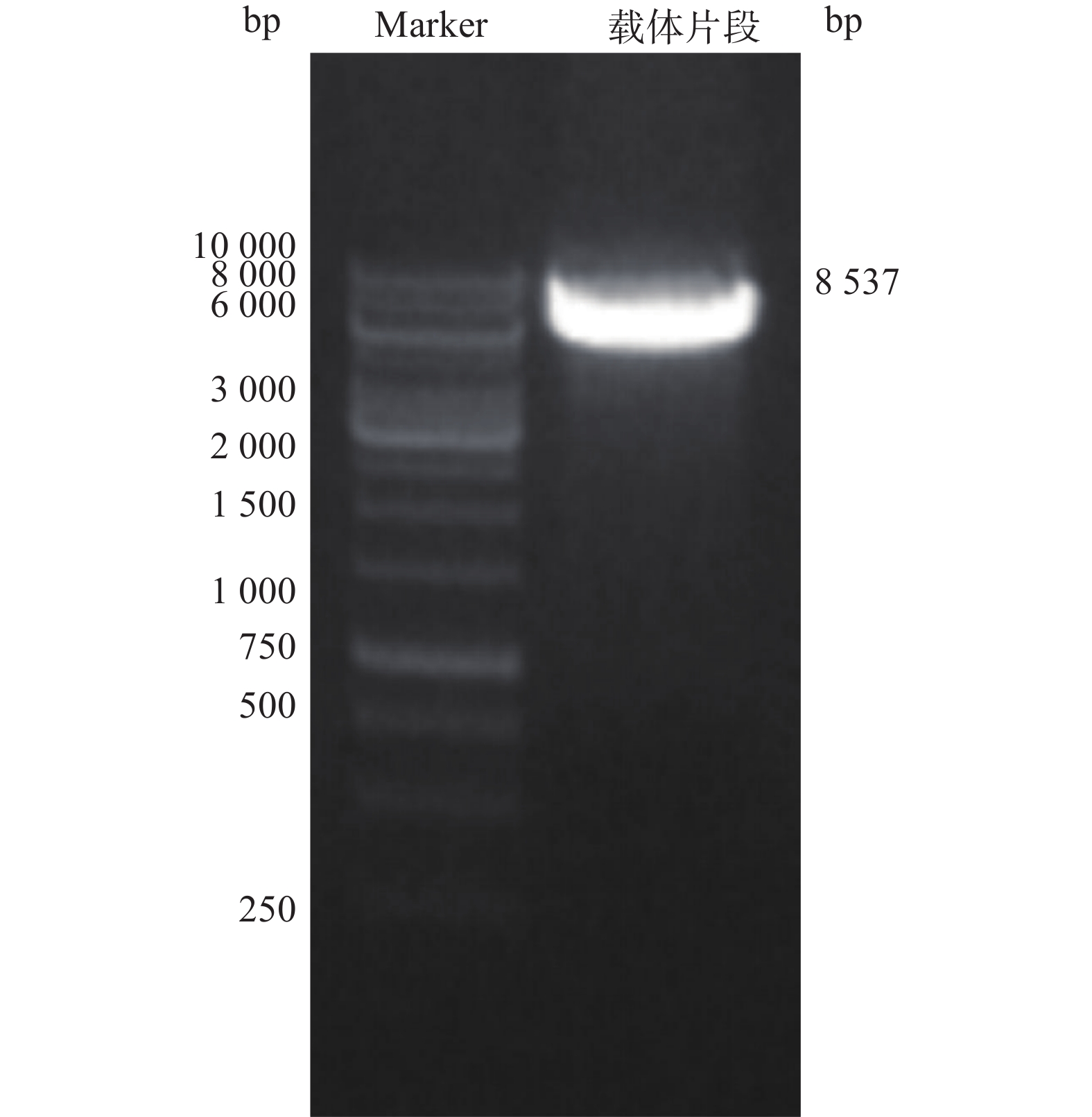
2.1.2 载体线性化、重组质粒的构建与鉴定
用限制性内切酶BamHI和XhoI酶切pMT406,胶回收线性化载体(大小约8 537 bp)。线性化的载体、PCR扩增的ZNF24与3FLAG产物,通过同源重组(无缝克隆)反应,将10 μl反应产物转化至DH5α。平皿培养过夜,挑取单克隆进行菌落PCR鉴定, 阳性克隆产物预期为1 438 bp。PCR阳性的克隆进一步测序鉴定,测序正确的重组质粒命名为pMT-ZNF24。
2.2 慢病毒载体包装、滴度测定
转染前24 h,胰酶消化并重悬293T细胞,取10个10 cm的培养皿,以1×107个/皿的细胞密度铺板。细胞贴壁后将原有培养基更换为Opti-MEM®培养基,体积为9 ml。取100 μg pMT-ZNF24(或pMT406)、65 μg pCMV-dR8.9、35 μg pCMV-VSV-G和适量Opti-MEM®培养基加入至15 ml无菌离心管中混匀,总体积为5 ml。再取500 μl细胞转染液和4.5 ml Opti-MEM®培养基混匀后滴加至上述离心管中,轻柔摇晃至均匀,室温孵育20 min。孵育完成后,将混合液分装到293T细胞中,每皿1 ml,轻轻摇晃混匀后放回培养箱。细胞培养6 h后弃上清液,加入10 ml DMEM培养基继续培养,2 d后收集细胞上清液。用60 ml 0.22 μm PVDF过滤装置过滤上清液, 4 ℃,25 000 r/min离心2 h,然后分装保存于−80 ℃冰箱。采用孔稀释法测定病毒滴度:准备5个EP管,各加入90 μl含10%FBS的高糖DMEM。EP管1中添加10 μl的待测病毒原液,EP管2中添加EP管1混合液10 μl,依次操作至EP管5。293T细胞接种到96孔板的 5个孔中,每孔约5×104个细胞,待细胞贴壁后去掉原液,依次加入EP管中的病毒液继续培养,24 h后换液,观察并记录3 d后稀释率最大孔中的荧光细胞数量。病毒滴度=荧光细胞数/病毒原液量。
2.3 慢病毒转染HCT116细胞
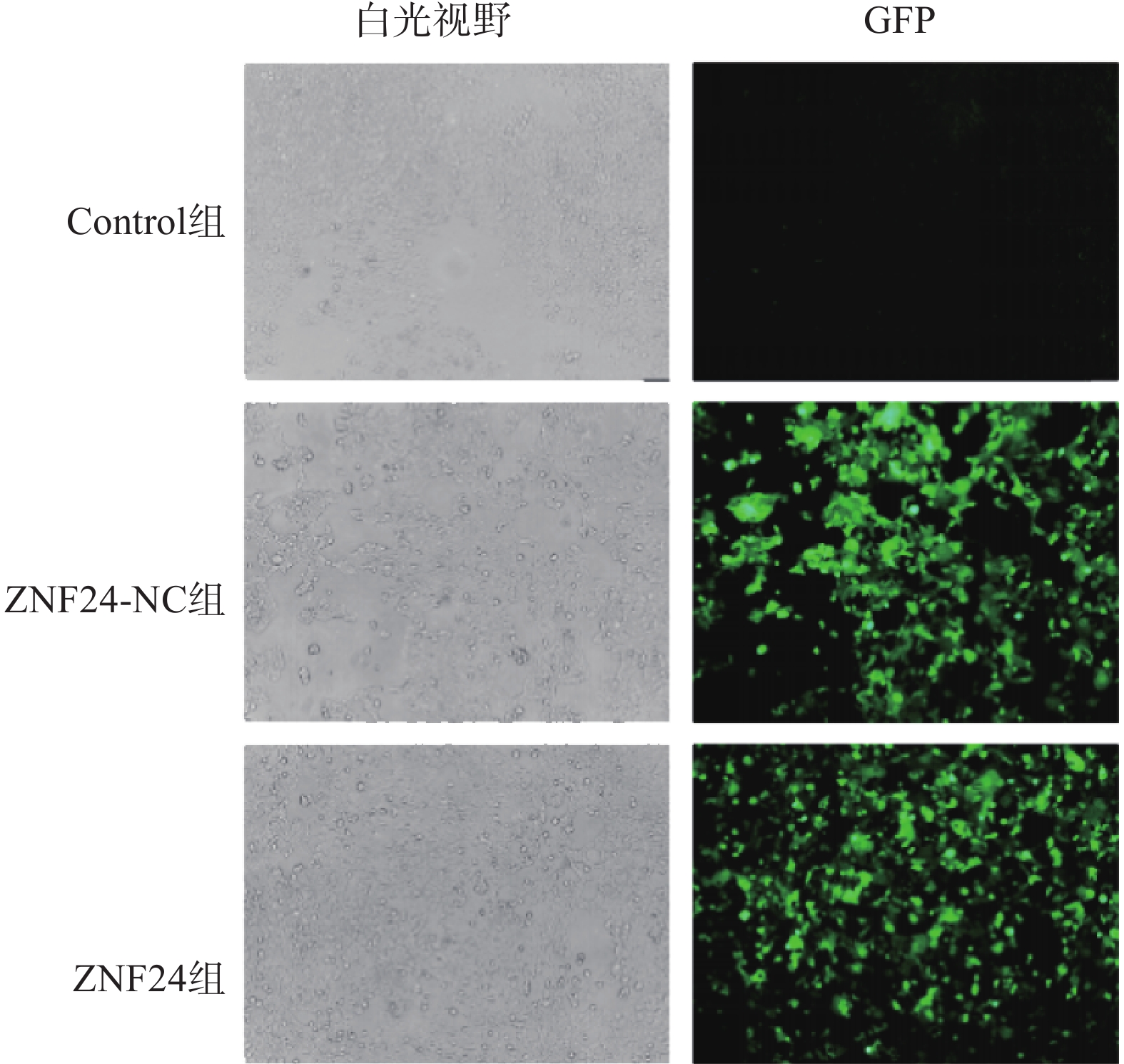
胰酶消化HCT116细胞,重悬后接种于24孔板中,每孔细胞约为3×105个,待细胞融合达30%时以细胞感染指数(MOI)=10计算病毒浓缩液体积并转染细胞。将HCT116细胞分为3组:空白对照组(HCT116细胞不转染病毒)、阴性对照组(HCT116细胞转染不含ZNF24的空载体慢病毒)和ZNF24组(HCT116细胞转染ZNF24过表达慢病毒)。转染72 h后,在ZNF24组和阴性对照组中加入 4 μg / ml嘌呤霉素,继续培养72 h后得到稳定表达细胞株。
2.4 实时荧光定量PCR检测ZNF24 mRNA的表达
慢病毒转染HCT116细胞,TRIzol法裂解细胞并提取RNA,mRNA反转录成cDNA后扩增ZNF24。ZNF24引物序列上游为CATTCCCTAAGGCACTGTGAT,下游为TTGAGGAACACCCATACTGAGA;GAPDH引物序列上游为TGACTTCAACAGCGACACCCA,下游为CACCCTGTTGCTGTAGCCAAA。2−ΔΔCt法分析ZNF24 mRNA的表达量。
2.5 蛋白印迹法检测ZNF24蛋白的表达
慢病毒转染HCT116细胞,裂解液(含蛋白酶抑制剂)裂解细胞,提取总蛋白并测定浓度。SDS-PAGE电泳后转模,脱脂牛奶封闭1 h,室温一抗孵育3 h,室温荧光素标记二抗孵育2 h,Odyssey双色红外激光成像系统检测荧光信号。
2.6 统计学分析
实验数据以3个独立试验的(
$\bar{x} \text{±} s$ )表示,采用 GraphPad Prism 5.0软件中单因素方差分析或t检验进行分析。3. 结果
3.1 PCR扩增及电泳结果
电泳结果显示,分别获得了大小约1 128 bp的ZNF24扩增产物与大小约111 bp的3FLAG扩增产物(图1)。
3.2 载体pMT406线性化
电泳结果显示,得到大小约8 537 bp线性化载体条带(图2)。
3.3 重组慢病毒载体pMT-ZNF24菌落PCR与测序鉴定
重组质粒经PCR扩增,电泳结果显示,获得约1 438 bp大小的阳性克隆PCR产物条带(图3)。对PCR产物进行测序比对分析,结果与目标序列完全一致。
3.4 病毒滴度检测结果及荧光显微镜下绿色荧光表达
ZNF24过表达慢病毒的滴度为3.25×109 TU/ml,ZNF24-NC慢病毒的滴度为6.19×109 TU/ml。以MOI=10计算病毒体积并转染HCT116细胞,4 μg /ml 嘌吟霉素筛选,荧光显微镜下约85%的细胞呈现绿色荧光蛋白(GFP)表达(图4)。
3.5 HCT116细胞中ZNF24 mRNA相对表达量比较
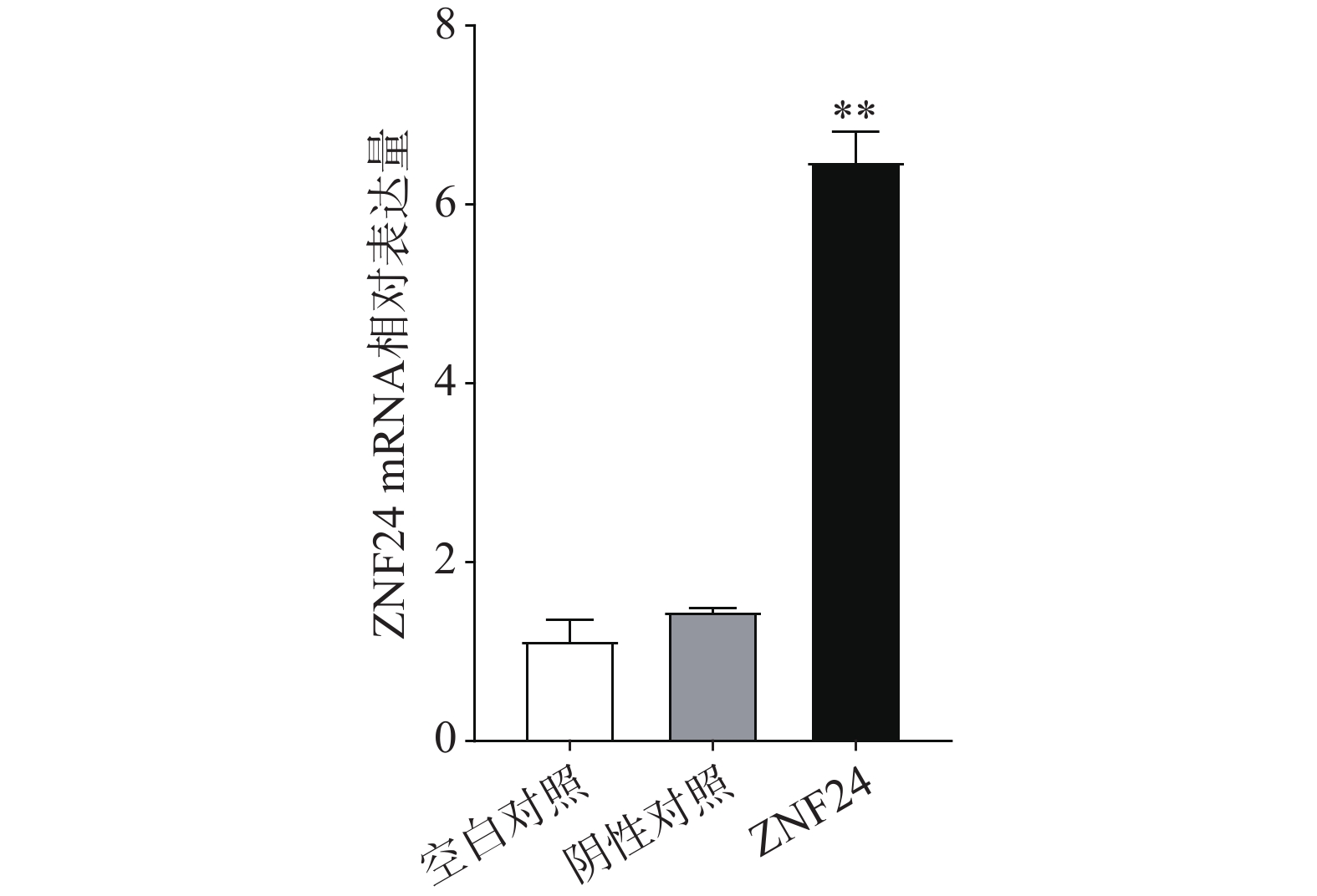
qRT-PCR结果表明,转染ZNF24过表达慢病毒的细胞组中ZNF24的mRNA表达显著高于阴性对照组和空白对照组(图5)。
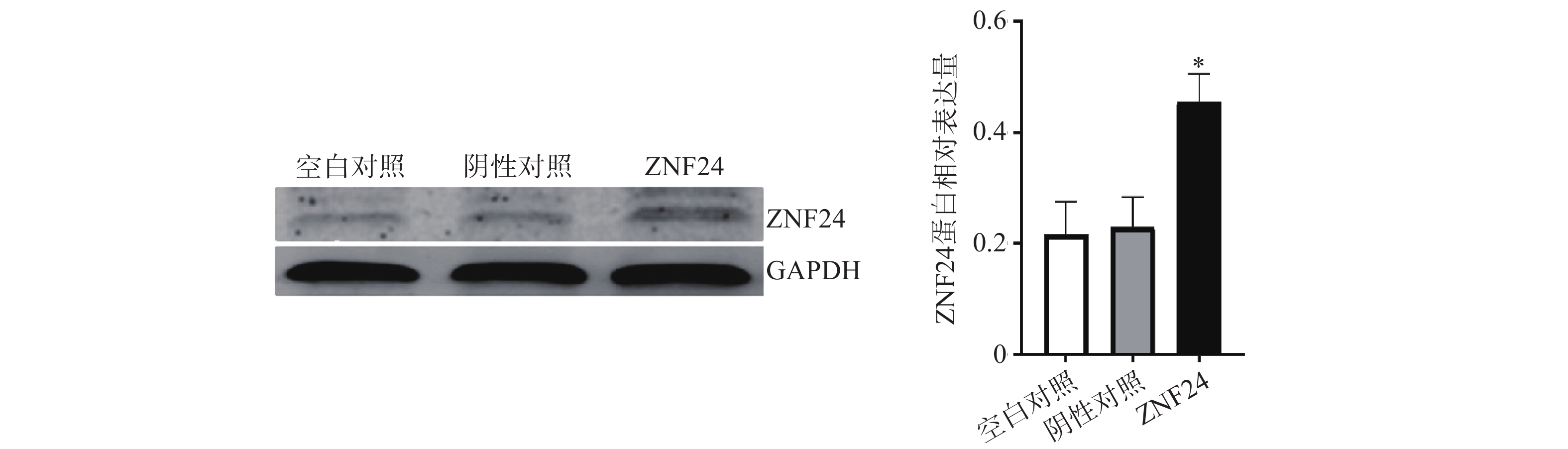
3.6 HCT116细胞中ZNF24蛋白相对表达量比较
蛋白印迹检测结果显示,转染ZNF24过表达慢病毒的细胞组中ZNF24表达水平明显高于阴性对照组和空白对照组 (图6)。
4. 讨论
结直肠癌是癌症致死的一个主要原因[16],致死的关键要素是其高水平的复发和转移[17]。结肠癌的发病机制十分复杂,涉及多种癌基因、抑癌基因的异常表达。因此,研究结直肠癌的发病机制,特别是结直肠癌复发转移的机制极其重要。研究表明,多种转录因子在结直肠癌等肿瘤中异常表达,参与结直肠癌的发病、转移和侵袭,如在肿瘤微环境参与VEGF表达调控的 STAT3转录因子已成为新的抗肿瘤药物的作用靶点[18]。
体外与乳腺癌细胞肿瘤动物模型的研究表明,ZNF24通过与血管内皮生长因子(VEGF)启动子上游序列(−144/−134, 非(TCAT) n重复序列)直接结合抑制VEGF基因转录,从而抑制血管增生达到抑制肿瘤生长[19]。然而,敲减人微血管内皮细胞中的ZNF24导致细胞迁移,侵袭和增殖减弱,暗示ZNF24具有促进人微血管内皮细胞的血管生成潜力[3]。多项研究表明ZNF24通过调控不同靶基因(如Twist1[11]、β-连环蛋白[8]和DGL1[10]等)的转录表达以及竞争性结合蛋白因子[8],在多种不同肿瘤的发生发展、侵袭和转移中起着重要复杂的两面性作用(促进或抑制)。
本研究成功构建了ZNF24过表达慢病毒载体,获得相应的病毒。转染HCT116细胞,获得稳定过表达ZNF24的HCT116细胞株,为开展后续研究提供了物质基础。
-
表 1 中药治疗系统性红斑狼疮的机制和靶点
药名 组成成分 作用机制/靶点 相关分子 单药 柴胡多糖 巨噬细胞 NO、TNF-α、IL-1β、IL-6 黑灵芝多糖 巨噬细胞 TLR4 雷公藤素 树突细胞 TLR9 淫羊藿 Treg/Th17 STAT5b 白芍总苷 Treg/Th17 IL-17 mRNA 青蒿素 B细胞 TLR7/9 复方 青蒿-鳖甲药 青蒿、鳖甲 脂质代谢 L-a-溶血卵磷脂 解毒祛瘀滋阴方 白花蛇草、紫草、赤芍、生地、炙鳖甲 氨基酸代谢 二十碳五烯酸、甘氨酸、
苯丙氨酸、色氨酸细胞凋亡 Bcl-2 清热化瘀滋阴方 麦冬、赤芍、水牛角片、炒白术、连翘、牡丹皮、
紫草、秦艽、青风藤、金银花、白花蛇草细胞凋亡 Bcl-2、bax、fas、p53、c-myc 解毒祛瘀滋肾方 生地黄、炙鳖甲、青蒿、白花蛇草、积雪草、赤芍、
牡丹皮、佛手、升麻、重楼、生甘草Treg CD299、CD25 凉血化瘀方 水牛角、生地黄、赤芍、牡丹皮、制大黄、玄参、知母 细胞因子 IL-6、IL-17、IL-21 益气养阴方 生薏仁、丹皮、芍药、白花蛇草、升麻、水牛角、丹参、青蒿、鳖甲 细胞因子 IL-6、IL-8、TNF-α 养阴清热煎 生地黄、牡丹皮、青蒿、白花蛇草、蛇莓、半枝莲、益母草、丹参 细胞黏附分子 ICAM-1、ICAM-3 滋阴清热方 山茱萸、生地黄、茯苓、泽泻、牡丹皮、青蒿、甘草 细胞凋亡 IL-1β、NLRP3 芪加真武汤 茯苓、白芍、附子、生姜、甘草、黄芪、刺五加 细胞凋亡 fas 中药组方 生地、黄芩、积雪草、川穹、丹参、猫抓草 细胞因子 IL-2、IL-4、IL-10 狼疮静 生地、熟地、丹皮、白花蛇草 细胞凋亡、B细胞 核酸抗原 狼疮定 白花蛇草、赤芍、生地、水牛角等 细胞因子 IL-2 狼疮方 白花蛇草、半枝莲、紫草、丹参、益母草 细胞因子 IL-6、IL-10 狼疮颗粒 生地黄、熟地黄、牡丹皮、白花蛇草 Treg/Th17 sIL-2R、TNF-α 昆仙胶囊 昆明山海棠、淫羊藿、枸杞子、菟丝子等 能量代谢 支链氨基酸、丙氨酸、糖蛋白、柠檬酸、磷酸胆碱、牛磺酸、葡萄糖 注:NLRP. 核苷酸结合寡聚化结构域样受体蛋白;IL. 白介素;ICAM. 细胞黏附分子;Treg. 调节性T细胞;Th17. T辅助细胞17;TNF. 肿瘤坏死因子;TLR. Toll 样受体  下载: 导出CSV
下载: 导出CSV
-
[1] LI M, ZHANG W, LENG X, et al. Chinese SLE Treatment and Research group (CSTAR) registry: I. Major clinical characteristics of Chinese patients with systemic lupus erythematosus[J]. Lupus,2013,22(11):1192-1199. doi: 10.1177/0961203313499086 [2] FELDMAN C H, MARTY F M, WINKELMAYER W C, et al. Comparative rates of serious infections among patients with systemic lupus erythematosus receiving immunosuppressive medications[J]. Arthritis Rheumatol,2017,69(2):387-397. doi: 10.1002/art.39849 [3] MORAND E F, FURIE R, TANAKA Y, et al. Trial of anifrolumab in active systemic lupus erythematosus[J]. N Engl J Med,2020,382(3):211-221. doi: 10.1056/NEJMoa1912196 [4] LEE H K, KIM E Y, KIM H S, et al. Effect of human mesenchymal stem cells on xenogeneic T and B cells isolated from lupus-prone MRL. Faslpr mice[J]. Stem Cells Int,2020,2020:5617192. [5] WANG D D, NIU L Y, FENG X B, et al. Long-term safety of umbilical cord mesenchymal stem cells transplantation for systemic lupus erythematosus: a 6-year follow-up study[J]. Clin Exp Med,2017,17(3):333-340. doi: 10.1007/s10238-016-0427-0 [6] DÖRNER T, FURIE R. Novel paradigms in systemic lupus erythematosus[J]. Lancet,2019,393(10188):2344-2358. doi: 10.1016/S0140-6736(19)30546-X [7] 潘杰. 系统性红斑狼疮中医证型与实验室指标及病变系统的相关性分析[D]. 南京: 南京中医药大学, 2015. [8] 苏励, 姚重华, 茅建春, 等. 风湿病 “同病异治” 病证相关性的代谢组学研究[C] //全国第八届中西医结合风湿病学术会议论文汇编, 广州, 2010: 25-29. [9] 周燕妮, 汤水福. 热毒血瘀型 LN 肾系膜细胞的蛋白质组学研究[C] //中国中西医结合学会肾脏疾病专业委员会2018年学术年会论文摘要汇编, 重庆, 2018: 1421. [10] 刘瑞萍, 朱妮, 李达, 等. 系统性红斑狼疮中医辨证分型与自身抗体谱的关系[J]. 广东医学, 2012, 33(2):273-275. doi: 10.3969/j.issn.1001-9448.2012.02.052 [11] 代荣琴, 潘磊, 杨丽娟. 系统性红斑狼疮患者自身抗体与中医辨证分型的关系[J]. 世界中西医结合杂志, 2016, 11(6):808-811. [12] 欧阳昕, 文锦丽, 彭武建, 等. 应用代谢组学方法构建系统性红斑狼疮诊断模型[J]. 中国现代医学杂志, 2011, 21(23):2878-2881,2885. doi: 10.3969/j.issn.1005-8982.2011.23.015 [13] 闻向晖, 范永升, 温成平. 系统性红斑狼疮证候特征、中医临床疗效及作用机制研究[J]. 浙江中医药大学学报, 2019, 43(10):1108-1113. doi: 10.16466/j.issn1005-5509.2019.10.020 [14] 鲁莹. 昆仙胶囊对MRL/lpr小鼠狼疮性肾炎代谢组学影响的研究[D]. 广州: 南方医科大学, 2011. [15] 陈娟, 邓军, 周佳, 等. 青蒿-鳖甲药对配伍治疗系统性红斑狼疮小鼠的代谢组学研究[J]. 中国药理学通报, 2016, 32(5):727-732. doi: 10.3969/j.issn.1001-1978.2016.05.026 [16] 余强. 黑灵芝多糖的免疫调节活性及其作用机制[D]. 南昌: 南昌大学, 2014. [17] 张丽萍. 雷公藤治疗系统性红斑狼疮的临床观察[J]. 中国现代药物应用, 2011, 5(19):85-86. doi: 10.3969/j.issn.1673-9523.2011.19.071 [18] 王卓龙. 白藜芦醇对Pristane诱导狼疮鼠作用研究[D]. 北京: 北京协和医学院, 2012. [19] 廖洁月. 淫羊藿素对系统性红斑狼疮Foxp3/IL17a调控作用与分子机制及其治疗MRL/1pr狼疮鼠的实验研究[D]. 长沙: 中南大学, 2014. [20] 刘喜德, 金实. 狼疮静颗粒对活动性系统性红斑狼疮患者血清可溶性白介素-2受体、肿瘤坏死因子的影响[J]. 中医杂志, 2003, 44(5):360-361. doi: 10.3321/j.issn:1001-1668.2003.05.024 [21] 陈雷鸣, 朱正阳, 范永升, 等. 解毒祛瘀滋肾方对MRL/lpr狼疮小鼠CD4+T细胞CD229/CD25轴的影响[J]. 中华中医药杂志, 2021, 36(4):1873-1877. [22] 郭云龙. 白芍总苷对MRL/lpr狼疮小鼠临床疗效及外周血IL-17、Foxp3 mRNA表达的影响[D]. 广州: 南方医科大学, 2013. [23] 简练. 白芍总苷对系统性红斑狼疮样小鼠的治疗作用及对Th17/Treg细胞免疫失衡的影响[D]. 广州: 南方医科大学, 2013. [24] 常杰, 李忱, 张文. 青蒿、青蒿素及其衍生物治疗系统性红斑狼疮研究现状[J]. 中国中西医结合杂志, 2019, 39(3):377-380. doi: 10.7661/j.cjim.20180521.084 [25] 吴言为. 青蒿素衍生物SM934对系统性红斑狼疮的疗效及作用机制研究[D]. 上海: 中国科学院上海药物研究所, 2016. [26] 温成平, 范永升, 李学铭. 中药狼疮定对系统性红斑狼疮患者血清可溶性白细胞介素-2受体和新喋呤水平的影响[J]. 中国中西医结合杂志, 2001, 21(5):339-341. doi: 10.3321/j.issn:1003-5370.2001.05.006 [27] 梁鸣, 李幼姬, 阳晓, 等. 中药狼疮方对狼疮样小鼠脾细胞体外分泌白细胞介素-6和白细胞介素-10的影响[J]. 中国中西医结合杂志, 2002, 22(5):372-375. doi: 10.3321/j.issn:1003-5370.2002.05.015 [28] 堵建岗, 沈建国, 王卫兰, 等. 中药组方对阴虚内热型系统性红斑狼疮的疗效及对Th1/Th2平衡的调节作用[J]. 中华中医药学刊, 2016, 34(7):1756-1758. doi: 10.13193/j.issn.1673-7717.2016.07.064 [29] 朱震, 余黎, 汪悦. 凉血化瘀方对MRL/lpr小鼠自身抗体及血清白细胞介素-6、17、21表达的影响[J]. 广州中医药大学学报, 2015, 32(6):1055-1058. [30] 田昱平, 彭飞, 陈涛, 等. 益气养阴活血方对系统性红斑狼疮患者血清炎症因子、生活质量及疗效的影响[J]. 广西医科大学学报, 2017, 34(9):1341-1343. [31] 谢晓辉, 杜晨光, 丁培杰, 等. 滋阴降火法对阴虚内热证系统性红斑狼疮患者细胞因子IL-10、IL-18的影响[J]. 实用临床医药杂志, 2017, 21(17):61-63. doi: 10.7619/jcmp.201717018 [32] 陈爱平, 熊佩华, 李福民, 等. 养阴清热煎剂对系统性红斑狼疮患者黏附分子表达及狼疮损伤指数的影响[J]. 中医杂志, 2005, 46(6):433-435. doi: 10.3321/j.issn:1001-1668.2005.06.013 [33] 黄钢花, 刘叶, 陈银环, 等. 清养透解法对自发性红斑狼疮模型小鼠血管损伤因子的影响[J]. 广州中医药大学学报, 2012, 29(5):555-557,611. [34] 陈明, 汤郁, 曾瑜, 等. 芪加真武汤对系统性红斑狼疮患者外周血淋巴细胞凋亡及Fas和FasL表达的影响[J]. 中国皮肤性病学杂志, 2005, 19(6):338-340. doi: 10.3969/j.issn.1001-7089.2005.06.007 [35] 胡晓琳. 中药干预对气阴两虚型系统性红斑狼疮PBMC凋亡及相关指标的影响[J]. 新中医, 2015, 47(6):99-101. [36] 温成平, 范永升, 许志良, 等. 解毒祛瘀滋阴药与激素对系统性红斑狼疮T细胞亚群Bcl-2基因表达的干预作用研究[J]. 中国中西医结合肾病杂志, 2006, 7(5):272-274. doi: 10.3969/j.issn.1009-587X.2006.05.008 [37] 刘喜德, 金实. 中药狼疮静颗粒对狼疮性BXSB小鼠脾脏CD4+T、CD19+B细胞凋亡的影响[J]. 中国中西医结合杂志, 2003, 23(9):692-694. doi: 10.3321/j.issn:1003-5370.2003.09.014 [38] 范萍, 梁康礼, 胡小倩, 等. 滋阴清热方对系统性红斑狼疮小鼠细胞焦亡相关因子的影响[J]. 中医杂志, 2017, 58(16):1404-1407. -

